
Биохимические показатели при поражении мышечной системы. Креатинурия. Что обеспечивает мышечное сокращение атф жиры вода минеральные соли
Механизмы мышечного сокращения
При световой микроскопии было замечено, что в момент сокращения ширина А-диска не уменьшается, а I-диски и Н-зоны саркомеров суживаются. При электронной, микроскопии было установлено, что длина нитей актина и миозина в момент соскращения не изменяется. Поэтому Хаксли и Хэнсон разработали теорию скольжения нитей. Согласно этой теории мышца укорачивается в результате движения тонких актиновых нитей в промежутки между миозиновыми. Это приводит к укорочению каждого саркомера, образующего миофибриллы. Скольжение же нитей обусловлено тем, что при переходе в активное состояние головки отростков миозина связываются с центрами актиновых нитей и вызывают их движение относительно себя (гребковые движения). Но это последний этап всего сократительного механизма.
Сокращение начинается с того, что в области концевой пластинки двигательного нерва возникает ПД. Он с большой скоростью распространяется по сарколемме и переходит с неё по, системе поперечных трубочек СР, на продольные трубочки и цистерны. Возникает деполяризация мембраны цистерн и из них в саркоплазму высвобождаются ионы кальция. На нитях актина расположены молекулы еще двух белков – тропонина и тропомиозина. При низкой (менее 10-8 моль) концентрации кальция, т.е. в состоянии покоя, тропомиозин блокирует присоединение мостиков миозина к нитям актина. Когда ионы кальция начинают выходить из СР, молекула тропонина изменяет свою форму таким образом, что освобождает активные центры актина от тропомиозина. К этим центрам присоединяются головки миозина и начинается скольжение за счет ритмического прикрепления и разъединения поперечных мостиков с нитями актина. При этом головки ритмически продвигаются по нитям актина к Z-мембранам. Для полного сокращения мышцы необходимо 50 таких циклов.
Передача сигнала от возбужденной мембраны к миофибриллам называется электромеханическим сопряжением. Когда генерация ПД прекращается и мембранный потенциал возвращается к исходному уровню, начинает работать Са-насос (фермент Са-АТФаза). Ионы кальция вновь закачиваются в цистерны саркоплазматического ретикулума и их концентрация падает ниже 10-8 моль. Молекулы тропонина приобретают исходную форму и тропомиозин вновь начинает блокировать активные центры актина. Головки миозина отсоединяются от них и мышца за счет эластичности приходит в исходное расслабленное состояние.
Энергетика мышечного сокращения
Источником энергии для сокращения и расслабления служит АТФ. На головках миозина есть каталитические центры, расщепляющие АТФ до АДФ и неорганического фосфата. Т.е. миозин является одновременно ферментом АТФ-азой. Активность миозина как АТФ-азы значительно возрастает при его взаимодействии с актином. При каждом цикле взаимодействия актина с головкой миозином расщепляется 1 молекула АТФ. Следовательно, чем больше мостиков переходят в активное состояние, тем больше расщепляется АТФ, тем сильнее сокращение. Для стимуляции АТФ-азной активности миозина требуются ионы кальция, выделяющиеся из саркоплазматического ретикулома (СР), которые способствуют освобождению активных центров актина от тропомиозина.
Однако запасы АТФ в клетке ограничены. Поэтому для восполнения запасов АТФ происходит его восстановление – ресинтез. Он осуществляется анаэробным и аэробным путем. Процесс анаэробного ресинтеза осуществляется фосфогенной и гликолитической системами. Фосфогенная система использует для восстановления АТФ запасы креатинфосфата. Он расщепляется на креатин и фосфат, который с помощью ферментов переносится на АДФ (АДФ+Фн=АТФ). Фосфогенная система ресинтеза обеспечивает наибольшую мощность сокращения, но в связи с малым количеством креатинфосфата в клетке, она функционирует лишь 5-6 секунд сокращения. Гликолитическая система использует для ресинтеза АТФ анаэробное расщепление глюкозы (гликогена) до молочной кислоты. Каждая молекула глюкозы обеспечивает восстановление трех молекул АТФ. Энергетические возможности этой системы выше, чем фосфагенной, но и она может служить источником энергии сокращения лишь 0,5-2 мин. При этом работа гликолитической системы сопровождается накоплением в мышцах молочной кислоты и снижением содержания кислорода. При продолжительной работе, с усилением кровообращения, ресинтез АТФ начинает осуществляться с помощью окислительного фосфорилирования, т.е. аэробным путем. Энергетические возможности окислительной системы значительно больше остальных. Процесс происходит за счет окисления углеводов и жиров. При интенсивной работе в основном окисляются углеводы, при умеренной – жиры. Для расслабления также нужна энергия АТФ. После смерти содержание АТФ в клетках быстро снижается и когда становится ниже критического, поперечные мостики миозина не могут отсоединиться от актиновых нитей (до ферментативного аутолиза этих белков). Возникает трупное окоченение. АТФ необходима для расслабления потому, что обеспечивает работу кальциевого насоса.
studfiles.net
Роль АТФ в механизмах мышечного сокращения. Энергетика мышечного сокращения
Энергия АТФ используется во время деятельности скелетной мышцы для 3-х процессов:
■ работы K+-Na+-насоса, обеспечивающего постоянство градиента концентраций ионов K+ и Na+ по обе стороны мембраны;
■ процесса скольжения актиновых и миозиновых нитей, ведущего к укорочению миофибрилл;
■ работы кальциевого насоса, необходимого для расслабления волокна.
При работе мышц химическая энергия превращается в механическую, т.е. мышца является химическим двигателем, а не тепловым. Для процессов сокращения и расслабления мышц потребляется энергия АТФ.Расщепление АТФ с отсоединением одной молекулы фосфата и образованием аденозиндифосфата (АДФ) сопровождается выделением 10 ккал энергии на 1 моль: АТФ = АДФ + Ф + Эн. Однако запасы АТФ в мышцах невелики (около 5 ммоль/л ). Их хватает лишь на 1 - 2 сработы. Количество АТФ в мышцах не может изменяться, т.к. при отсутствии АТФ в мышцах развивается контрактура (не работает кальциевый насос и мышцы не в состоянии расслабляться), а при избытке - теряется эластичность.
Для продолжения работы требуется постоянное восполнение запасов АТФ.Восстановление АТФ происходит в анаэробных условиях- за счет распада креатинфосфата (КрФ) и глюкозы (реакции гликолиза), в аэробных условиях- за счет реакций окисления жиров и углеводов.
Быстрое восстановление АТФпроисходит в тысячные доли секунды за счет распада КрФ: АДФ + КрФ = АТФ + Кр. Наибольшей эффективности этот путь энергообразования достигает к 5 - 6-й секундеработы, но затем запасы КрФ исчерпываются, т.к. их также немного (около 30 ммоль/л).
Медленное восстановление АТФ в анаэробных условиях обеспечивается энергией расщепления глюкозы (выделяемой из гликогена) – реакцией гликолиза с образованием в конечном итоге молочной кислоты (лак-тата) и восстановлением двух молекул АТФ. Эта реакция достигает наибольшей мощности к концу 1-й минуты работы. Особое значение этот путь энергообразования имеет при высокой мощности работы, которая продолжается от 20с до 1 – 2мин (например, при беге на средние дистанции), а также при резком увеличении мощности более длительной и менее мощной работы (финишные ускорения при беге на длинные дистанции) и при недостатке кислорода во время выполнения статической работы.Ограничение использования углеводов связано не с уменьшением запасов гликогена (глюкозы) в мышцах и в печени, а с угнетением реакции гликолиза избытком накопившейся в мышцах молочной кислоты.
Реакции окисления обеспечивают энергией работу мышц в условиях достаточного поступления в организм кислорода, т.е. при аэробной работе длительностью более 2 – 3мин.Доставка кислорода достигает необходимого уровня после достаточного развертывания функций кислородтранспортных систем организма (дыхательной, сердечно-сосудистой систем и системы крови). Важным показателем мощности аэробных процессов является предельная величина поступления в организм кислорода за 1 мин - максимальное потребление кислорода (МПК).Эта величина зависит от индивидуальных возможностей каждого человека. У нетренированных лиц в 1 мин поступает к работающим мышцам около 2,5 – 3 л О2, а у высококвалифицированных спортсменов (лыжников, пловцов, бегунов-стайеров и др.) достигает 5 – 6 л и даже 7 л в 1 мин.
При значительной мощности работы и огромной потребности при этом в кислороде основным субстратом окисления в большинстве спортивных упражнений являются углеводы,т.к. для их окисления требуется гораздо меньше кислорода, чем при окислении жиров. При использовании одной молекулы глюкозы (С6Н12О6), полученной из гликогена, образуется 38 молекул АТФ, т.е. аэробный путь энергообразования обеспечивает при том же расходе углеводов во много раз больше продукции АТФ, чем анаэробный путь. Молочная кислота в этих реакциях не накапливается, а промежуточный продукт – пировиноградная кислота – сразу окисляется до конечных продуктов обмена – СО2 и Н2О.
В качестве источника энергии жиры используются в состоянии двигательного покоя, при любой работе сравнительно невысокой мощности (требующей до 50 % МПК) и при очень длительной работе на выносливость (требующей около 70 – 80 % МПК). Среди всех источников энергии жиры обладают наибольшей энергетической емкостью:при расходовании 1 моля АТФ выделяется около 10 ккал энергии, 1 моля КрФ – око-ло 10,5 ккал, 1 моля глюкозы при анаэробном расщеплении – около 50 ккал, а при окислении 1 моля глюкозы в аэробных условиях – около 700 ккал, при окислении 1 моля жиров – 2 400 ккал. Однако использование жиров при работах высокой мощности лимитируется трудностью доставки кислорода работающим тканям.
Работа мышц сопровождается выделением тепла. Теплообразова-ние происходит в момент сокращения мышц – начальное теплообразование (оно составляет всего одну тысячную всех энерготрат) и в период восстановления – запаздывающее теплообразование.
В обычных условиях при работе мышц тепловые потери составляют около 80 % всех энерготрат. Для оценки эффективности механической работы мышцы используют вычисление коэффициента полезного действия (кпд). Величина кпд показывает, какая часть затрачиваемой энергии используется на выполнение механической работы мышцы. Ее вычисляют по формуле
кпд = [А : (Е - е)] · 100 %,
где А – энергия, затраченная на полезную работу;
Е – общий расход энергии;
е – расход энергии в состоянии покоя за время, равное длительности работы.
У нетренированного человека кпд примерно 20 %, у спортсмена – 30 – 35 %, т.е. мышца использует на движение 20 – 35 % химической энергии, остальная часть в форме тепла передается кровью другим тканям и равномерно согревает организм. Вот почему на холоде человек старается больше двигаться – подогревает себя энергией мышц. Мелкие непроизвольные сокращения мышц вызывают дрожь – организм увеличивает образование тепла.
При ходьбе наибольший кпд отмечается при скорости 3,6 – 4,8 км/ ч, при педалировании на велоэргометре – при длительности цикла около 1 сек. С увеличением мощности работы и включением «ненужных» мышц кпд уменьшается. При статической работе, поскольку А = 0, эффективность работы оценивается по длительности поддерживаемого напряжения мышц.
Трехглавая плеча
Наружная бедра
Камбаловидная


 67 % —
67 % —
33 %
49 %
51 %
16 %
Рис. 24. Состав мышечных волокон в разных мышцах:
медленные; ,.*>%' - быстрые
V<>-
Материалы для самостоятельной подготовки
Вопросы к коллоквиуму и для самоконтроля
1. Какие виды мышц у позвоночных животных и человека Вы знаете?
2. Назовите функции скелетных мышц.
3. Перечислите нейроны, иннервирующие скелетные мышцы.
4. Что является функциональной единицей мышцы?
5. Что входит в состав двигательной единицы (ДЕ)?
6. Что называют мотонейронным пулом?
7. Дать характеристику больших и малых ДЕ.
8. В чем заключается правило Хеннемана?
9. Опишите структуру мышечного волокна.
10. Как устроены миофибриллы?
11. Что такое саркомер?
12. Чем можно объяснить, что в состоянии покоя мышца имеет поперечнополо-сатый вид в световом микроскопе?
13. Опишите строение актиновых и миозиновых нитей.
14. Какова роль потенциала действия в возникновении мышечного сокращения?
15. Опишите механизм сокращения, расслабления мышечного волокна.
16. Кем была открыта ферментативная активность миозина?
17. Укажите последовательность событий, ведущих к сокращению, а затем рас-слаблению мышечного волокна.
18. В чем заключается роль АТФ в механизмах мышечного сокращения?
19. Перечислите фазы одиночного сокращения мышцы.
20. В каких случаях происходит суммация сокращений? Что такое тетанус?
21. Какие формы тетануса Вы знаете?
22. От чего зависит сокращение целой мышцы?
23. В чем заключается метод электромиографии?
24. От каких факторов зависит амплитуда ЭМГ?
25. Что такое сила мышцы и от каких морфологических и физиологических факторов она зависит?
26. Перечислите типы мышечных волокон. Дайте их характеристику.
27. Назовите режимы работы мышц.
28. Опишите энергетику мышечного сокращения.
infopedia.su
Биохимические механизмы сокращения и расслабления мышц
Биохимический цикл мышечного сокращения состоит из 5 стадий:
1 стадия – в стадии покоя миозиновая «головка» может гидролизовать АТФ до АДФ и Фн, но не обеспечивает освобождения продуктов гидролиза. Образуется стабильный комплекс: миозин-АДФ-Фн.
2 стадия – возбуждение двигательного нерва приводит к освобождению ионов Са2+ из саркоплазматического ритикулума мышечного волокна. Ионы Са2+ связываются тропонином С (Тн-С). В результате этого взаимодействия изменяется конформация всей молекулы тропонина, а затем – тропомиозина. Вследствие этого в актине открываются центры связывания с миозином. Миозиновая «головка» связывается с F-актином, образуя с осью фибриллы угол около 900.
3 стадия – присоединение актина к миозину обеспечивает высвобождение АДФ и Фн из актин-миозинового комплекса. Это приводит к изменению конформации этого комплекса и угол между актином и миозиновой «головкой» изменяется с 900 до 450. В результате изменения угла филаменты актина втягиваются между филаментами миозина, т. е. происходит их скольжение навстречу друг другу. Укорачиваются саркомеры, сокращаются мышечные волокна.
4 стадия – новая молекула АТФ связывается с комплексом актин-миозин.
5 стадия – комплекс миозин-АТФ обладает низким сродством к актину и поэтому происходит отделение миозиновой «головки» от F-актина. Филаменты возвращаются в исходное состояние, мышца расслабляется. Затем цикл возобновляется.
Н2О
актин
АТФ-миозин
5 1
актин-миозин-АТФ миозин-АДФ-Фн
2
АТФ актин
3
актин-миозин актин-миозин-АДФ-Фн
АДФ, Фн
Рис. 33.1. Цикл мышечного сокращения
Движущая сила мышечного сокращения – энергия, освобождающаяся при гидролизе АТФ.
Роль ионов кальция в регуляции мышечного сокращения
Ключевая роль в регуляции мышечного сокращения принадлежит ионам кальция (Са2+). Миофибриллы обладают способностью взаимодействовать с АТФ и сокращаться лишь при наличии в среде определенных концентраций ионов кальция. В покоящейся мышце концентрация ионов Са2+ поддерживается ниже пороговой величины при участии Са2+-зависимой АТФазы. В состоянии покоя эта система активного транспорта накапливает кальций в цистернах саркоплазматического ретикулума и трубочках Т-системы.
Мышечное сокращение инициируется приходом потенциала действия на концевую пластинку двигательного нерва. В синапс выделяется ацетилхолин, который связывается с постсинаптическими рецепторами мышечного волокна. Далее потенциал действия распространяется вдоль сарколеммы к поперечным трубочкам Т-системы и происходит передача сигнала на цистерны саркоплазматического ретикулума. Последние начинают освобождать находящийся в них кальций в саркоплазму. Концентрация Са2+ увеличивается с 10-7 до 10-5 ммоль/л. Кальций связывается с Тн-С, что вызывает конформационные сдвиги, передающиеся на тропомиозин и далее на актин. Открываются закрытые ранее центры в актине для связывания с миозином. Актин взаимодействует с миозином, что инициирует сокращение мышечного волокна.
После прекращения действия двигательного импульса кальций с помощью Са2+-зависимой АТФазы откачивается из цитоплазмы в цистерны саркоплазматического ретикулума. Уход кальция из комплекса с Тн-С приводит к смещению тропомиозина и закрытию активных центров актина. Миозиновая «головка» отсоединяется от актина. Мышца расслабляется.
Кальций является аллостерическим модулятором мышечного сокращения.
studfiles.net
Энергетика мышечного сокращения
Сокращение – изменение механического состояния миофибриллярного сократительного аппарата мышечных волокон в результате дейтвия нервных импульсов. Скелетная мышца представляет собой сложную систему, преобразующую химическую энергию в механическую работу и тепло.
По теории скольжения, в основе сокращения лежит механическое взаимодействие между миозиновыми и актиновыми миофиламентами благодоря образованию между ними в период активности попереречных мостиков.
Непосредственным источником энергии для мышечного сокращения является расщепление высокоэнергетического вещества АТФ. В мышце происходит также промежуточная реакция, вовлекающая 2-ое высокоэнергетическое вещество – креатинфосфат (КФ). Оно не может действовать как непосредственный источник энергии, поскольку его расщепление не оказывает влияние на сократительные белки мышцы. КФ обеспечивает энергией ресинтез АТФ. В свою очередь, энергия для ресинтеза КФ обеспечивается окислением.
Молекулярный механизм сокращения мышечного волокна состоит в том, что возникающий на мембране в области концевой пластинки потенциал действия распространяется по системе поперечных трубочек вглубь волокна, вызывает деполяризацию мембран цистерн саркоплазматического ретикулума и освобождение из них ионов кальция. Свободные ионы кальция в межфибриллярном пространстве запускают процесс сокращения.
Совокупность процессов, обуславливающих распространение потенциала действия вглубь мышечного волокна, выход ионов кальция из саркоплазматического ретикулума, взаимодействие сократительных белков и укорочение мышечного волокна называют "электрическим сопряжением". Энергия гребкового движения одного мостика производит перемещение на 1% длины актиновой нити. Для дальнейшего скольжения сократительных белков друг относительно друга мостики между актином и миозином должны распадаться и вновь образовываться на следующем центре связывания Са2+-. Такой процесс происходит в результате активации в этот момент молекул миозина. Миозин приобретает свойства фермента АТФ-азы, который вызывает распад АТФ. Выделившаяся при распаде АТФ энергия приводит к разрушению имеющихся мостиков и образованию в присутствии Са2+новых мостиков на следующем участке актиновой нити. В результате повторения подобных процессов многократного образования и распада мостиков сокращается длина отдельных саркомеров и всего мышечного волокна в целом.
Максимальная концентрация кальция в миофибрилле достигается уже через 3 мс после появления потенциала действия в поперечных трубочках, а максимальное напряжение мышечного волокна - через 20 мс.
Весь процесс от появления мышечного потенциала действия до сокращения мышечного волокна называется электромеханической связью (или электромеханическим сопряжением). В результате сокращения мышечного волокна актин и миозин более равномерно распределяются внутри саркомера, и исчезает видимая под микроскопом поперечная исчерченность мышцы.
Основа сокращения мышцы — биохимические процессы, которые совершаются в 2 фазы: первую, анаэробную (бескислородную), и вторую, аэробную (кислородную). В каждой из этих фаз происходит расщепление веществ с освобождением энергии и их восстановление (ресинтез). Поэтому мышца, лишенная кислорода, может долго работать при условии удаления остаточных продуктов обмена веществ. Следует учесть, что исключительная роль в биохимических процессах первой фазы принадлежит кислоторастворимым фосфорным соединениям, относительное содержание которых в скелетных мышцах кролика равняется (в процентах кислоторастворимого фосфора): аденозинтрифосфорная кислота (АТФ) — 35, креатинфосфорная кислота — 45, гексозомонофосфорная кислота — 5, неорганическая фосфорная кислота — 10.
Первое место в анаэробной фазе химических процессов занимает расщепление АТФ, которое происходит в первую очередь и с наибольшей скоростью. АТФ дефосфорилируется благодаря ферментативному действию мышечных белков. Отщепляя группу фосфорной кислоты, АТФ превращается сначала в аденозиидифосфорную (АДФ), а затем при потере еще одной группы фосфорной кислоты — в адениловую кислоту. При отщеплении каждого моля фосфорной кислоты освобождается 33,5 — 50,2 кДж, а в результате отщепления двух молей фосфорной кислоты — до 100,5 кДж при превращении каждого моля АТФ в адениловую кислоту. Эта энергия используется для сокращения мышц. Так как АТФ расходуется, то длительная мышечная работа невозможна без ресинтеза АТФ.
Ресинтез АТФ в анаэробных условиях происходит за счет энергии, которая освобождается при втором, более медленном процессе дефосфорилироваиия креатинфосфорной кислоты на креатин и фосфорную кислоту. При этом расщеплении на каждый моль креатинфосфата освобождается 46 кДж. Часть креатина необратимо распадается.
И, наконец, происходит третий, наиболее медленный процесс анаэробной фазы — расщепление гексозофосфорной кислоты на фосфорную и молочную кислоты. При этом процессе на каждый моль образовавшейся молочной кислоты выделяется 104,6 кДж Гликоген, присоединяя фосфорную кислоту, сначала превращается в гексозомонофосфат, а затем в гексозодифосфат с небольшим поглощением энергии. Гексозодифосфат распадается на молочную и фосфорную кислоты с значительным освобождением энергии.
Ресинтез АТФ совершается в течение тысячных долей секунды в результате присоединения к адениловой кислоте групп фосфорной кислоты, освободившихся при расщеплении креатинфосфорной кислоты и гексозофосфата, Ресинтез креатинфосфорной кислоты происходит в результате присоединения к креатину фосфорной кислоты, освободившейся при расщеплении гексозофосфата. Энергия, освободившаяся при расщеплении креатинфосфорной кислоты и гексозофосфата, обеспечивает процессы ресинтеза, главным образом АТФ. Так как избыток адениловой кислоты, образовавшейся в первой фазе из АТФ, необратимо дезаминируется и превращается в инозиновую кислоту и аммиак, то при полном ресинтезе АТФ вначале идет ресинтез адениловой кислоты. Адениловая кислота ресинтезируется из инозиновой кислоты и аммиака, освобождающегося при дезаминировании аминокислот.
Во второй, аэробной, фазе биохимических процессов освобождается наибольшее количество энергии, которое используется как для ресинтеза соединений, расщепляющихся в первой и второй фазах, так и главным образом для мышечной деятельности. В результате гликолиза (бескислородного распада глюкозы на две частицы молочной кислоты) и расщепления гексозофосфатов образуется молочная кислота, которая окисляется до углекислоты и воды. Но не вся молочная кислота окисляется, а примерно 76 часть (при утомлении около 1/4). При этом расщепление каждого моля молочной кислоты освобождает 1465,5 кДж. За счет этой энергии происходит ресинтез остальной части молочной кислоты (5/6 – 3/4) до глюкозы и гликогена, а также ресинтез АТФ и креатинфосфорной кислоты.
Следовательно, наибольшее количество энергии при мышечной работе освобождается при окислении углеводов. Гликоген содержится главным образом в анизотропных дисках. В мышцах в покое, а также при восстановлении после сокращения кроме углеводов расщепляется небольшая часть белков и липидов. При достаточном кровообращении и содержании глюкозы и кислорода в крови мышца работает за счет энергии окислительных процессов. В покое мышца использует около 5% кислорода, поступающего с кровью. Ресинтез веществ, расщепляющихся при мышечной деятельности, увеличивается при повышении интенсивности тканевого дыхания. Таким образом, процесс расщепления усиливает ресинтез, что ведет к наиболее экономному использованию веществ и энергии.
В результате сокращения в мышечных волокнах возникает напряжение. Эта способность развивать напряжение является основным свойством мышцы. Напряжение, развиваемое мышцами при сокращении, реализуется по-разному. Если внешняя нагрузка меньше, чем напряжение сокращающейся мышцы, то мышца укорачивается и вызывает движение – это изотонический (концентрический, миометрический) тип сокращения. Поскольку в экспериментальных условиях при электрическом раздражении укорочение мышцы происходит при постоянном напряжении = внешней нагрузке.
Если внешняя нагрузка больше, чем напряжение, развиваемое мышцей во время сокращения, то такая мышца растягивается при сокращении – это эксцентрический (плиометрический) тип сокращения. Изотонический и эксцентрический типы сокращения, т.е. сокращения. при которых мышца изменяет свою длину, относятся к динамической форме сокращения.
Сокращение мышцы, при котором она развивает напряжение, но не изменяет своей длины, называется изометрическим. Это статическая форма сокращения. Она возникает в 2-ух случаях: либо когда внешняя нагрузка = напряжению, развиваемому мышцей при сокращении, либо когда внешняя нагрузка превышает напряжение мышцы, но отсутствуют условия для растяжения мышцы под влиянием этой внешней нагрузки.
При динамических формах сокращения производится внешняя работа – при изотоническом сокращении положительная, при эксцентрическом отрицательная. Величина работы в обоих случаях может быть определена как произведение внешней нагрузки (поднятого веса) на пройденное расстояние. При изометрическом сокращении "расстояние" = 0, и, согласно физическому закону, в этом случае мышца не производит никакой работы.
Если внешняя нагрузка на мышцы меньше, чем ее напряжение, мышца укорачивается и вызывает движение. Это концентрический, или миометрический, тип сокращения. В экспериментальных условиях при электрическом раздражении изолированной мышцы ее укорочение происходит при постоянном напряжении, равном внешней нагрузке. Поэтому этот тип сокращения называют также изотоническим (изос - равный, тонус - напряжение).
Если внешняя нагрузка на мышцы больше, чем ее напряжение, развиваемое во время сокращения, мышца растягивается (удлиняется). Это эксцентрический, или плиометрический, тип сокращения. Концентрический и эксцентрический типы сокращения, т.е. сокращения, при которых мышца изменяет длину, относятся к динамической форме сокращения.
Сокращение мышцы, при котором она развивает напряжение, но не изменяет своей длины, называется изометрическим (изос - равный, метр - длина). Это статическая форма сокращения. Она возникает в двух случаях: когда внешняя нагрузка равна напряжению, развиваемому мышцей при сокращении, либо когда внешняя нагрузка превышает напряжение мышцы, но отсутствуют условия для растяжения мышцы под влиянием этой внешней нагрузки.
При динамических формах сокращения производится внешняя работа: при концентрическом сокращении - положительная, при эксцентрическом - отрицательная. Величина работы в обоих случаях определяется как произведение внешней нагрузки (поднятого веса) на пройденное расстояние. При изометрическом сокращении "расстояние" равно нулю, и, согласно физическому закону, в этом случае мышца не производит никакой работы. Однако с физиологической точки зрения изометрическое сокращение требует расхода энергии и может быть очень утомительным. В этом случае работа может быть определена как произведение величины напряжения мышцы на время ее сокращения (т.е. эквивалентно импульсу силы в физике). Во время изометрического сокращения в тепло превращается вся выделяемая мышцей энергия, а динамическом сокращении не менее 50% ее энергии.
Энергетические резервы человека ( с массой тела 75 кг)
| Источники энергии | Энергоемкость, кДж | Возможная продолжительность работы, с |
| АТФ | 4 - 5 | 2 - 3 |
| Креатинфосфат (КрФ) | 14 - 15 | 15 - 20 |
| Гликоген+глюкоза | 4600 - 13000 | 120 - 240 |
| Жиры | 300000 - 400000 | более 240 |
biofile.ru
Химический состав мышечной ткани | Бесплатные курсовые, рефераты и дипломные работы
В мышечной ткани человека содержится 72–80% воды и 20–28% сухого остатка от массы мышцы. Вода входит в состав большинства клеточных структур и служит растворителем для многих веществ. Большую часть сухого остатка образуют белки и другие органические соединения (табл. 2).

Рис. 4. Схема расположения Т-систем и саркоплазматического ретикулума в мышечном волокне
Таблица 2
Химический состав скелетных мышц млекопитающих
| Компонент | % на сырую массу | Компонент | % на сырую массу |
| Вода | … 72–80 | АТФ | 0,25–0,40 |
| Сухой остаток: | 20–28 | карнозин | 0,20–0,30 |
| белки | 16,50–20,90 | карнитин | 0,02–0,05 |
| гликоген | 0,30–3,00 | ансерин | 0,09–0,15 |
| фосфолипиды | 0,40–1,00 | свободные аминокислоты | 0,10–0,70 |
| холестерин | 0,06–0,20 | молочная кислота | 0,01–0,02 |
| креатинфосфат | 0,20–0,55 | зола | 1,00–1,50 |
| креатин | 0,003–0,005 |
Основные белки мышц
Среди белков мышечной ткани выделяют три основные группы: саркоплазматические белки, на долю которых приходится около 35%, миофибриллярные белки, составляющие около 45%, и белки стромы, количество которых достигает 20%.
Саркоплазматические белки растворимы в воде и слабых солевых растворах. Основную массу их составляют белки-ферменты, локализованные главным образом в митохондриях и катализирующие процессы окислительного фосфорилирования, а также многие ферменты гликолиза, азотистого и липидного обменов, находящиеся в саркоплазме. К этой группе относится также белок миоглобин, который связывает кислород с большим сродством, чем гемоглобин, и депонирует молекулярный кислород в мышцах. В последнее время открыта группа саркоплазматических белков парвальбуминов, которые способны связывать ионы кальция, однако их физиологическая роль остается не выясненой.
Миофибриллярные белки включают сократительные белки миозин, актин и актомиозин, а также регуляторные белки тропомиозин, тропонин, α- и β-актинины. Миофибриллярные белки обеспечивают сократительную функцию мышц.
Миозин является одним из основных сократительных белков мышц и составляет около 55% общего количества мышечных белков. Из него состоят толстые нити (филаменты) миофибрилл. Молекулярная масса этого белка – около 470 000. В молекуле миозина различают длинную фибриллярную часть и глобулярные структуры (головки). Фибриллярная часть молекулы миозина имеет двуспиральную структуру (рис. 5). В составе молекулы выделяют шесть субъединиц: две тяжелые полипептидные цепи (молекулярная масса 200 000) и четыре легкие цепи (молекулярная масса 1500–2700), расположенные в глобулярной части. Основной функцией фибриллярной части молекулы миозина является способность образовывать хорошо упорядоченные пучки миозиновых филаментов или толстые протофибриллы (см. рис. 5). На головках молекулы миозина расположены активный центр АТФ-азы и актинсвязывающий центр, поэтому они обеспечивают гидролиз АТФ и взаимодействие с актиновыми филаментами.

Рис. 5. Схема строения молекулы миозина (а), миозинового пучка (б) и миозиновой толстой нити (в)
Молекула миозина содержит значительное количество глутаминовой аминокислоты и имеет большой отрицательный заряд, что усиливает связывание свободных ионов Са2+ и Мg2+. В присутствии ионов Са2+ повышается АТФ-азная активность миозина и скорость гидролиза АТФ согласно уравнению
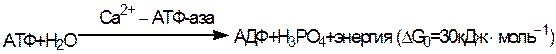
Химическая энергия АТФ, освобождающаяся в ходе данной ферментативной реакции, используется для изменения конформации белка миозина и генерации напряжения между толстыми и тонкими нитями миозина в сокращающейся мышце. Посредством ионов Mg2+ миозин способен присоединять молекулы АТФ и АДФ, а также взаимодействовать с молекулами актина, находящимися в составе тонких нитей миофибрилл.
Актин – второй сократительный белок мышц, который составляет основу тонких нитей (рис. 6). Известны две его формы – глобулярный G-актин и фибриллярный F-актин. Глобулярный актин – это шарообразный белок с молекулярной массой 42 000. На его долю приходится около 25% общей массы мышечного белка. В присутствии Мg2+ актин подвергается нековалентной полимеризации с образованием нерастворимого филамента в виде спирали, получившего название F-актин. Обе формы актина не обладают ферментативной активностью. Каждая молекула G-актина способна связывать один ион Ca2+, который играет важную роль в инициировании сокращения. Кроме того, молекула G-актина прочно связывает одну молекулу АТФ или АДФ. Связывание АТФ G-актином обычно сопровождается его полимеризацией с образованием F-актина и одновременным расщеплением АТФ до АДФ и фосфата. АДФ остается связанной с фибриллярным актином. Процесс полимеризации актина можно описать следующим уравнением:

F-актин активирует АТФ-азу миозина, что создает движущую силу процессу сокращения.

Рис. 6.Схема строения актиновой, или тонкой нити
Актин способен взаимодействовать с миозином, образуя актомиозиновый комплекс. Молярное соотношение актина и миозина в актомиозиновом комплексе – примерно 1:1. Нить F-актина может связывать большое число молекул миозина. Существенным свойством актомиозинового комплекса является диссоциация его в присутствии АТФ и Мg2+.
В состав тонких нитей наряду с актином входят и другие минорные белки – тропомиозин, тропонины, актинины.
Тропомиозин (Тм) – это структурный белок актиновой нити, представляющий собой вытянутую в виде тяжа молекулу. Две его полипептидные цепи как бы обвивают актиновые нити (см. рис. 6). На концах каждой молекулы тропомиозина расположены белки тропониновой системы, наличие которой характерно только для поперечно-полосатых мышц.
Тропонин (Тн) является регуляторным белком актиновой нити. Он состоит из трех субъединиц – ТнТ, ТнI и ТнС. Тропонин Т(ТнТ) обеспечивает связывание этих белков с тропомиозином. Тропонин I (ТнI) блокирует (ингибирует) взаимодействие актина с миозином. Тропонин С (ТнС) – это Са2+-связывающий белок, структура и функции которого подобны широко распространенному в природе белку кальмодулину. Тропонин С, как и кальмодулин, связывает четыре иона Са2+ на молекулу белка и имеет молекулярную массу 17 000. В присутствии Са2+ изменяется конформация тропонина С, что приводит к изменению положения Тн по отношению к актину, в результате чего открывается центр взаимодействия актина с миозином.
Таким образом, тонкий филамент миофибриллы поперечно-полосатой мышцы состоит из F-актина, тропомиозина и трех тропониновых компонентов – ТнС, ТнI и ТнТ. Кроме этих белков, в мышечном сокращении участвует белок актинин. Обнаруживается он в зоне Z-линии, к которой крепятся концы F-актиновых молекул тонких нитей миофибрилл.
Белки мышечной стромы в скелетной мышце представлены в основном коллагеном и эластином, которые входят в состав сарколеммы и Z-линий миофибрилл. Эти белки обладают эластичностью, большой упругостью, что имеет существенное значение для процесса сокращения и расслабления мышцы.
Небелковые компоненты мышц
В состав сухого остатка мышц наряду с белками входят и другие вещества, среди которых выделяют азотсодержащие, безазотистые экстративные вещества и минеральные вещества.
К азотсодержащим веществам скелетных мышц относятся АТФ и продукты ее расщепления – АДФ и АМФ, а также креатинфосфат, креатин, креатинин, карнозин, ансерин, свободные аминокислоты и др.
АТФ, содержащаяся в количестве 0,25–0,40%, и креатинфосфат, количество которого колеблется в пределах 0,4–1,0%, являются источниками энергии мышечного сокращения. Продукты их распада – АДФ, АМФ и креатин – оказывают регулирующее действие на обмен веществ в мышцах. Карнозин является дипептидом и участвует в переносе фосфатных групп, стимулирует работу ионных насосов, увеличивает амплитуду мышечного сокращения, которые снижаются при утомлении и этим способствуют восстановлению работоспособности. Карнитин участвует в переносе жирных кислот – важных энергетических источников – через мембраны митохондрий и тем самым способствует их окислению и энергообразованию.
В состав различных клеточных мембран мышечной ткани входит ряд азотсодержащих фосфолипидов: фосфатидилхолин (лецитин), фосфатидилэтаноламин (кефалин), фосфатидилсерин и др. фосфолипиды участвуют в обменных процессах, являясь поставщиками холина и жирных кислот – субстратов тканевого дыхания. Другие азотсодержащие вещества – мочевина, мочевая кислота, пуриновые основания (аденин, гуанин) – являются промежуточными или конечными продуктами азотистого обмена и встречаются в мышцах в небольших количествах.
К безазотистым соединениям мышечной ткани относится гликоген, который находится в саркоплазме в свободном или связанном с белками состоянии и используется в мышцах как основной энергетический субстрат при напряженной работе. Количество его в зависимости от пищевого рациона питания и степени тренированности колеблется от 0,3 до 3,0% общей массы мышц. При тренировке увеличивается главным образом количество свободного гликогена. В мышце содержится ряд промежуточных продуктов обмена углеводов – гексозофосфаты, пировиноградная и молочная кислоты.
Из липидов в мышечной ткани обнаруживаются триглицериды в виде капелек жира, а также холестерин.
Минеральные вещества составляют 1–1,5% общей массы мышцы. Состав их разнообразен. Основными катионами являются Na+, К+, Mg2+, Са2+. Катионы К+ сосредоточены в основном внутри мышечных волокон, а Na+ – преимущественно в межклеточной жидкости. Они участвуют в процессах возбуждения мышц, запуске их сокращения. Среди анионов преобладают Сl–, Н2РО4–, НРО42–, SO42–, НСО3–.
В мышечной ткани содержится ряд микроэлементов: кобальт, железо, никель, бор, цинк и др. Они являются либо структурными компонентами сложных белковых молекул, либо активаторами ферментов. Все минеральные вещества играют важную роль в регуляции различных биохимических процессов в мышцах.
refac.ru
Энергетика мышечного сокращения. Биомеханика мышечных сокращений
Энергетика мышечного сокращения
Источником энергии для сокращения и расслабления служит АТФ. На головке миозина есть каталитические центры, расщепляющие АТФ до АДФ и неорганического фосфата, т.е. миозин является одновременно ферментом АТФ-азой. Активность миозина как АТФ-азы, значительно возрастает при его взаимодействии с актином. При каждом цикле взаимодействия актина с головкой миозина расщепляется 1 молекула АТФ. Следовательно, чем больше мостиков переходит в актиновое состояние, тем больше расщепляется АТФ, тем сильнее сокращения. Для стимуляции АТФ-азной активности миозина требуются ионы кальция, выделяющиеся из саркоплазматического ретикулума, которые способствуют освобождению активных центров актина от тропомиозина.

Мышцы человека
Однако запасы АТФ в клетке ограничены. Поэтому для восполнения запасов АТФ происходит ее восстановление – ресинтез. Он осуществляется анаэробным и аэробным путями. Процесс анаэробного ресинтеза осуществляется фосфогенной и гликолитической системами. Фосфогенная система использует для восстановления АТФ запасы креатинфосфата. Он расщепляется на креатин и фосфат, который с помощью ферментов переносится на АДФ (АДФЗ + ФН = АТФ). Фосфагенная система ресинтеза обеспечивает наибольшую мощность сокращения, но в связи с малым количеством креатинфосфата в клетке, она функционирует лишь 5-6 секунд сокращения. Гликолитическая система использует для ресинтеза АТФ анаэробное расщепление глюкозы (гликогена) до молочной кислоты. Каждая молекула глюкозы обеспечивает восстановление трех молекул АТФ. Энергетические возможности этой системы выше, чем фосфогенной, но и она может служить источником энергии сокращения лишь 0,5-2 минуты. При этом работа гликолитической системы сопровождается накоплением в мышцах молочной кислоты и снижением содержания кислорода. При продолжительной работе, с усилением кровообращения, ресинтез АТФ начинает осуществляться с помощью окислительного фосфорилирования, т.е. аэробным путем. Энергетические возможности окислительной системы значительно больше остальных. Процесс происходит за счет окисления углеводов и жиров. При интенсивной работе в основном окисляется углерод, при умеренной – жиры. Для расслабления также нужна энергия АТФ. После смерти содержание АТФ в клетках быстро снижается и когда становится ниже критической, поперечные мостики миозина не могут отсоединиться от актиновых нитей (до ферментативного аутолизма этих белков). Возникает трупное окоченение. АТФ необходима для расслабления потому, что обеспечивает работу кальциевого насоса.
Биомеханика мышечных сокращений
При нанесении на двигательный нерв или мышцу одиночного порогового или сверхпорогового раздражения, возникает одиночное сокращение. При его графической регистрации, на полученной кривой можно выделить три последовательных периода:
1. Латентный период. Это время от момента нанесения раздражения до начала сокращения. Его длительность около 1-2 мс. Во время латентного периода генерируется и распространяется потенциал действия, происходит высвобождение кальция из саркоплазматического ретикулума, взаимодействие актина с миозином и т.д.
2. Период укорочения (сокращения). В зависимости от типа мышцы (быстрая или медленная) его продолжительность от10 до 100 мсек. Это период от начала сокращения до максимального сокращения.
3. Период расслабления. Его длительность несколько больше, чем укорочения.
В режиме одиночного сокращения мышца способна работать длительное время без утомления, но его сила незначительна. Поэтому в организме такие сокращения встречаются редко, например, так могут сокращаться быстрые глазодвигательные мышцы. Чаще одиночные сокращения суммируются.
Суммация – это сложение двух последовательных сокращений при нанесении на мышцу двух пороговых или сверхпороговых раздражений, интервал между которыми меньше длительности одиночного сокращения, но больше продолжительности рефракторного периода, т.е. два раздражения попадает в фазу экзальтации.
Различают 2 вида суммации: полную и неполную суммацию. Неполная суммация возникает в том случае, если повторное раздражение наносится на мышцу, когда она начинает расслабляться. Полное возникает тогда, когда повторное раздражение действует до начала периода расслабления, т.е. конец периода укорочения. Амплитуда сокращения при полной суммации выше, чем неполной. Если интервал между двумя раздражениями еще больше уменьшить, например, нанести второе в середине периода укорочения, то суммации не будет, потому что мышца находится в состоянии рефрактерности (относительная рефракторная фаза).
Тетанус – это длительное сокращение мышцы, возникающее в результате суммации нескольких одиночных сокращений, развивающихся при нанесении на нее ряда последовательных раздражений. Различают 2 формы тетануса: зубчатый и гладкий.
Зубчатый тетанус наблюдается в том случае, если каждое последующее раздражение действует на мышцу, когда она уже начала расслабляться, т.е. наблюдается неполная суммация. Гладкий тетанус возникает тогда, когда каждое последующее раздражение наносится в конце периода укорочения, т.е. имеет место полная суммация отдельных сокращений. Амплитуда гладкого тетануса больше, чем зубчатого. В норме мышцы человека сокращаются в режиме гладкого тетануса. Зубчатый возникает при патологии, например, тремор рук при алкогольной интоксикации и болезни Паркинсона.
biofile.ru
Биохимические показатели при поражении мышечной системы. Креатинурия.
Мышечный аппарат человека и животных характеризуется полифункциональностью. Однако основной
функцией мышц является осуществление двигательного акта, т.е. сокращение и расслабление. При сокращении
мышц осуществляется работа, связанная с превращением химической энергии в механическую. В данном
разделе в основном рассматривается структурная основа процесса сокращения поперечно-полосатых мышц
позвоночных, поскольку этот процесс изучен наиболее полно. Как отмечалось, сократительная система
поперечно-полосатой мышцы состоит из перекрывающихся белковых нитей, которые скользят относительно
друг друга. Сокращение происходит за счет энергии, освобождающейся пргидролизе АТФ. В поперечно-
полосатой мышце сокращение зависит от концентрации ионов Са2+, которая в свою очередь регулируется
саркоплазматическим ретикулумом – специализированной системой мембран, накапливающей Са2+ в состоянии
покоя и высвобожающей его при воз- действии на мышечное волокно нервного импульса. Источники энергии
мышечной деятельности. Прежде всего ресинтез АТФ обеспечивается трансфосфорилированием АДФ с
креатинфосфатом. Данная реакция катализируется ферментом креатинкиназой. Креатинкиназный путь ресинтеза
АТФ является чрезвычайно быстрым и максимально эффективным (за счет каждой молекулы креатинфосфата
образуется молекула АТФ). Именно поэтому долгое время не удавалось установить уменьшение концентрации
АТФ и соответственно повышение концентрации АДФ даже при достаточно продолжительном тетанусе.
Применив специфический ингибитор креатинкиназы (1-фтор-2,4-динитрофенол), а также с помощью агентов,
препятствующих окислительному фосфорилированию АДФ в АТФ, Т. При работе умеренной интенсивности
мышца может покрывать свои энергетические затраты за счет аэробного метаболизма. Однако при больших
нагрузках, когда возможность снабжения кислородом отстает от потребности в нем, мышца вынуждена
использовать гликолитический путь снабжения энергией. При интенсивной мышечной работе скорость
расщепления гликогена или глюкозы с образованием молочной кислоты увеличивается в сотни раз.
Соответственно содержание молочной кислоты в мышечной ткани может повышаться до 1,0–1,2 г/кг и более. С
током крови значительное количество молочной кислоты поступает в печень, где ресинтезируется в глюкозу и
гликоген (глюконеогенез) за счет энергии окислительных процессов. Перечисленные механизмы ресинтеза АТФ
при мышечной деятельности включаются в строго определенной последовательности. Наиболее экстренным
является креатинкиназный механизм, и лишь примерно через 20 с максимально интенсивной работы начинается
усиление гликолиза, интенсивность которого достигает максимума через 40–80 с. При более длительной, а
следовательно, и менее интенсивной работе все большее значение приобретает аэробный путь ресинтеза АТФ.
Содержание АТФ и креатинфосфата в сердечной мышце ниже, чем в скелетной мускулатуре, а расход АТФ
велик. В связи с этим ресинтез АТФ в миокарде должен происходить намного интенсивнее, чем в скелетной
мускулатуре. Для сердечной мышцы теплокровных животных и человека основным путем образования богатых
энергией фосфорных соединений является путь окислительного фосфорилирования, связанный с поглощением
кислорода. Регенерация АТФ в процессе анаэробного расщепления углеводов (гликолиз) в сердце человека
практического значения не имеет. Именно поэтому сердечная мышца очень чувствительна к недостатку
кислорода. Характерной особенностью обмена веществ в сердечной мышце по сравнению со скелетной является
также то, что аэробное окисление веществ неуглеводной природы при работе сердечной мышцы имеет большее
значение, чем при сокращении скелетной мышцы. Только 30–35% кислорода, поглощаемого сердцем в норме,
расходуется на окисление углеводов и продуктов их превращения. Главным субстратом дыхания в сердечной
мышце являются жирные кислоты. Окисление неуглеводных веществ обеспечивает около 65–70% потребности
миокарда в энергии. Из свободных жирных кислот всердечной мышце особенно легко подвергается окислению
олеиновая кислота.
| Количественное определение общих липидов в сыворотке крови. | Общие липиды крови – это ТАГ, жирные кислоты, ФЛ, холестерин, находящиеся в различных фракциях липопротеинов крови. После гидролиза серной кислотой с фосфованилиновым реактивом появляется красное окрашивание, интенсивность которого пропорциональна содержанию липидов и измеряется на ФЭКе. | 4 – 8 г/л | При патологии чаще наблюдается увеличение содержания общих липидов за счет той или иной фракции (гиперлипемия). Концентрация общих липидов крови увеличивается при ожирении, атеросклерозе, ИБС, при сахарном диабете, циррозе печени, липоидном циррозе, пенкреатите. |

ПЕРВЫЙ ВОПРОС ИСКАТЬ В ИНТЕРНЕТЕ
Водно-солевой обмен. Взаимосвязь воды и солей. Значение воды. Распределение ее в организме.
Потребность в воде и ее выведение. Физико-химические процессы регуляции транспорта жидкости в условиях
Целостного организма. Минеральные соли (катионы и анионы) и осмотическое давление. Значение отдельных
infopedia.su










