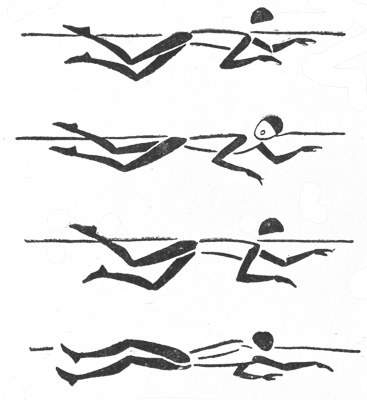
Окислительные пути распада углеводов в тканях. Анаэробное окисление углеводов
Анаэробное окисление углеводов
Анаэробное окисление углеводов происходит в клетках, органах и тканях без участия кислорода. Если процесс начинается с превращения глюкозы и заканчивается образованием молочной кислоты, то он называется анаэробным гликолизом, если начинается с превращения гликогена – гликогенолизом.
Гликолиз
На 1-й стадии гликолиза (рис. 7) происходит фосфорилирование глюкозы под действием фермента гексокиназы в присутствии АТФ и ионов Mg2+ с образованием глюкозо-6-фосфата (Г-6-Ф), который на 2-й стадии превращается во фруктозо-6-фосфат (Ф-6-Ф). Эту реакцию катализирует глюкозофосфатизомераза. В ходе 3-й стадии осуществляется фосфорилирование фруктозо-6-фосфата, в результате чего образуется фруктозо-1,6-дифосфат (Ф-1,6-Ф). Катализ данной реакции обеспечивает в присутствии ионов Mg2+ и АТФ фосфофруктокиназа.
На 4-й стадии Ф-1,6-Ф под действием альдолазы расщепляется на 2 фосфотриозы - глицеральдегид-3-фосфат (ГА-3-Ф) и дигидроксиацетонфосфат (ДАФ).
ДАФ под действием триозофосфатизомеразы превращается в ГА-3-Ф. Таким образом, из одной молекулы глюкозы мы получаем 2 молекулы ГА-3-Ф.
ГА-3-Ф в присутствии глицеральдегидфосфатдегидрогеназы, кофермента НАД+ и фосфорной кислоты окисляется с образованием 1,3-дифосфоглицерата (1,3-ДФГ) и НАДН(Н+). Энергия, высвобождающаяся в этой реакции аккумулируется в макроэргической связи 1,3-ДФГ, который далее превращается в 3-фосфоглицерат под действием фосфоглицераткиназы. Этот процесс сопряжен с фосфорилированием АДФ (образуется АТФ на уровне субстратного фосфорилирования).
3-ФГ при участии фосфоглицератмутазы превращается в 2-фосфоглицерат (2-ФГ), который подвергается дегидратации в присутствии енолазы и ионов Mg2+. В результате образуется фосфоенолпируват (ФЕП), содержащий макроэргическую связь. ФЕП под действием пируваткиназы при наличии ионов Mg2+ превращается в енольную форму пирувата. Этот процесс сопряжен с синтезом АТФ на уровне субстратного фосфорилирования. Енольная форма пирувата таутомеризуется в кетонную форму.
На заключительной стадии гликолиза происходит восстановление пирувата до L-лактата в присутствии НАДН(Н+) и фермента лактатдегидрогеназы. Образующийся в этой реакции кофермент НАД+ в дальнейшем используется в реакции окисления ГА-3-Ф.
Таким образом конечными продуктами гликолиза являются 2 молекулы L-лактата и 2 молекулы АТФ в расчете на 1 молекулу глюкозы.
Регуляция гликолиза осуществляется на уровне фосфофруктокиназной реакции. Фермент ингибируется высоким содержанием АТФ и цитрата. Аллостерическими активаторами фосфофруктокиназы являются АМФ, АДФ, Ф-6-Ф.
Гликогенолиз
Гликоген представляет собой разветвленный полисахарид, состоящий из остатков -D-глюкозы, связанных между собой в линейных участках молекулы -1,4-гликозидными связями, а в точках ветвления -1,6-гликозидными связями.
Под действием гликогенфосфорилазы происходит отщепление одного глюкозного остатка в линейных участках с переносом его на молекулу фосфорной кислоты, в результате чего образуется глюкозо-1-фосфат (Г-1-Ф). Гликогенфосфорилаза работает до тех пор, пока до ближайшей точки ветвления не останется 4 остатка глюкозы, затем в работу включается фермент олигосахаридтрансфераза, переносящий фрагмент из 3-х глюкозных остатков на соседнюю ветвь, таким образом, в точке ветвления остается один глюкозный остаток. Он отщепляется гидролитическим путем с помощью -1,6-гликозидазы в виде свободной молекулы глюкозы и вновь создаются условия для работы гликогенфосфорилазы в линейной цепи.
Продукт глигогенфосфорилазной реакции Г-1-Ф далее под действием фосфоглюкомутазы превращается в Г-6-Ф, который вовлекается в гликолитический путь (рис. 8).
Рис. 8. Схема распада гликогена в мышцах и печени.
Конечные продукты гликогенолиза – 2 молекулы L-лактата и 3 молекулы АТФ в расчете на 1 молекулу глюкозы.
studfiles.net
Анаэробное окисление углеводов — МегаЛекции
Основным источником энергии для клеток животных, растений и многих бактерий, является окисление глюкозы, протекающее в две стадии: анаэробную и аэробную.
Анаэробная стадия окисления глюкозы в клетках человека и животных получила название гликолиз. Гликолиз является первым, а в анаэробных условиях основным процессом катаболизма глюкозы, освобождающаяся при этом энергия аккумулируется в молекулах АТФ. Гликолиз протекает в цитоплазме, состоит из 10 реакций и условно делится на три фазы.
Первая – активирование глюкозы (пусковая реакция) и подготовка к расщеплению на две фосфотриозы; расщепление фруктозо-1,6-дифосфата и изомеризация дигидроксиацетонфосфата в глицеральдегид-3-фосфат.
Глюкоза
АТФ
АДФ гексокиназа
глюкозо – 6 – фосфат
гексозофосфатизомераза
фруктозо – 6 – фосфат
АТФ фосфофруктокиназа
АДФ
фруктозо- 1,6 –дифосфат
альдолаза
дигидроксиацетонфосфат триозофосфат- глицеральдегид–3–фосфат
изомераза
(2) глицеральдегид-3-фосфат
2 НАД+ 2 НАД× Н2
2 Фнглицеральдегидфосфат-
дегидрогеназа
(2)1,3 – дифосфоглицериновая кислота
2АДФ
фосфоглицераткиназа
2 АТФ
(2) 3 – фосфоглицериновая кислота
фосфоглицеромутаза
(2) 2 – фосфоглицериновая кислота
енолаза
2 Н2О
(2) фосфоенолпировиноградная кислота
2 АДФ
пируваткиназа
2 АТФ
(2) пировиноградная кислота
Рис. 4. Схема гликолиза
В рамках помещены исходные субстраты и конечные продукты гликолиза. Цифрами в скобках обозначено число молекул
Вторая – окисление двух молекул глицеральдегид-3-фосфата до двух молекул пирувата и аккумуляция освобождающейся энергии в сопряженном процессе субстратного фосфорилирования АДФ с образованием АТФ.
Третья – в анаэробных условиях, для поддержания процесса, НАД+ выполняет роль кофермента-переносчика пары водорода от глицеральдегид-3-фосфата на пируват, обеспечивая клетку энергией.
Значение гликолиза особенно велико для тканей с ограниченным доступом кислорода и испытывающих периодически резкое возрастание потребления АТФ.
Гликолиз может начинаться либо с фосфорилирования глюкозы, либо с фосфоролиза гликогена (или крахмала в растениях). В скелетных мышцах оба пути выражены в равной степени, а в мышце сердца и головном мозге преобладает фосфорилирование глюкозы.
Брожение – это окисление органических веществ, в том числе и углеводов, различными микроорганизмами в анаэробных условиях с целью получения энергии. Брожение происходит в окружающей среде, в пищевых продуктах. В отраслях пищевой промышленности чаще всего используется молочнокислое и спиртовое брожение. В данных видах брожения окислению подвергается глюкоза, и их химизм до образования пировиноградной кислоты совпадает с гликолизом.
Молочнокислое брожение – это процесс получения энергии молочнокислыми бактериями. По характеру различают два вида молочнокислого брожения: гомоферментативное и гетероферментативное, которые осуществляются соответствующими группами молочнокислых бактерий. Гомоферментативные (однотипнобродящие бактерии), в процессе брожения образуют в основном молочную кислоту и очень мало побочных продуктов. Они окисляют углеводы молока по пути гликолиза в анаэробных условиях, где пируват служит акцептором водорода от НАД·Н2 и восстанавливается в конечный продукт брожения – лактат. Суммарное уравнение этого типа брожения можно записать так:
С6Н12О6 → 2СН3―СНОН―СООН + 2АТФ.
Лактат, накапливаясь до рН 4,8-4,6, вызывает скисание молока. Этот процесс лежит в основе квашения капусты, огурцов, помидор и других продуктов растительного происхождения, силосования кормов для животных. Образующийся лактат предотвращает развитие гнилостных бактерий, плесневых грибов, т.е. служит консервантом.
Гетероферментативные бактерии (разнотипнобродящие) – наряду с молочной кислотой образуют значительное количество других веществ: этанола, СО2, уксусной кислоты. Окисление углеводов молока гетероферментативными молочнокислыми бактериями осуществляется своеобразной ферментативной системой в которой нет фермента альдолазы, но есть ферменты пентозофосфатного цикла и других типов брожения. После фосфорилирования гексоза окисляется дегидрогеназой и декарбоксилируется, превращаясь в пентозофосфат. Последний расщепляется на глицеральдегид-3-фосфат и ацетилфосфат. Глицеральдегид-3-фосфат, как и у гомоферментативных молочнокислых бактерий, окисляется до пирувата, который затем восстанавливается в лактат, а НАД·Н2 окисляется в НАД+. Ацетилфосфат дефосфорилируется и превращается в ацетат (уксусную кислоту), частично восстанавливается (через уксусный альдегид) в этанол. Таким образом, конечными акцепторами водорода в этом типе брожения служат пируват и уксусный альдегид.
Культуры гетероферментативных молочнокислых бактерий используют в производстве кефира, кумыса, курунги, мацони и других продуктов.
Спиртовое брожение осуществляется клетками дрожжевых грибов для получения энергии в анаэробных условиях. Большинство дрожжей сбраживает моносахариды, а именно глюкозу по пути гликолиза до образования пирувата. В анаэробных условиях, фермент дрожжей пируватдекарбоксилаза, превращает пируват в уксусный альдегид. Последний восстанавливается ферментом алкогольдегидрогеназой в этанол с окислением НАД·Н2:
пируватдекарбоксилаза О
2 СН3―СО―СООН 2 СН3С + 2 СО2
пируват альдегид Н
О алкогольдегидрогеназа
2 СН3 ―С + 2 НАД×Н2 2 СН3―СН2ОН + 2 НАД+.
Н ЭТАНОЛ
Суммарное уравнение спиртового брожения:
С6Н12О6 → 2 С2Н5ОН + 2 СО2 + 2 АТФ.
Пропионовокислые бактерии сбраживают углеводы до пропионовой, уксусной кислот и углекислого газа. Одна молекула пирувата окисляется до уксусной кислоты и СО2 и две молекулы пирувата превращаются в пропионовую кислоту.
Уксуснокислые бактериии окисляют этанол и другие спирты до уксусной кислоты: + О2
СН3-СН2ОН СН3-СООН + Н2О
Этанол Ацетат
Есть микроорганизмы осуществляющие маслянокислое, лимоннокислое и другие виды брожения. Многие брожения используются в пищевых технологиях и не менее важную роль они выполняют в природе.
Анаэробную стадию окисления глюкозы наиболее удобно изучать на спиртовом брожении, которое протекает по пути гликолиза. Процесс состоит из 12 реакций.
ХОД РАБОТЫ. В ступку вносят 0,5-1 г дрожжей и растирают до однородной массы добавляя раствор с массовой долей сахарозы или глюкозы 5 %. Содержимое из ступки переносят в трубку Эйнгорна (рис.5), смывая несколько раз ступку и пестик 3-4 мл раствором сахаров так, чтобы запаянное колено было полностью заполнено смесью.
Трубку помещают в термостат при 37-38 °С на 2-3 часа. Интенсивность брожения учитывают каждые 15-30 мин по объему СО2, который собирается в запаянном колене (в см3). Если брожение протекает интенсивно, то после каждого замера запаянное колено трубки Эйнгорна повторно заполняют той же бродящей смесью (смесь перемещают из расширенной части трубки в запаянное колено). После 4-5 замеров строят график интенсивности брожения. По оси ординат откладывают объем СО2, по оси абсцисс – время замера (30, 60, 90, 120 и 150 мин от начала термостатирования).
ОБНАРУЖЕНИЕ ЭТАНОЛА. По окончании работы, отфильтровывают через складчатый фильтр 3-5 мл бродящей смеси в пробирку, добавляют 3-5 капель раствора Люголя и по каплям 10 % раствор гидроксида натрия до обесцвечивания раствора. После этого пробирку с раствором нагревают на кипящей водяной бане. В зависимости от количества образовавшегося этанола через 3-5 мин появляется запах хлороформа.
С2Н5ОН + 4 I2 + 6 NaOH СНI + 5NaI + HCOONa + 5 h3O
Этанол Иодоформ
ОБНАРУЖЕНИЕ УГЛЕКИСЛОГО ГАЗА. После последнего замера проделывают качественную реакцию на СО2. Для чего в бродильный прибор (не перемещая в нем смесь) приливают до краев раствор с массовой долей гидроксида натрия 10 %, закрывают отверстие большим пальцем и содержимое хорошо перемешивают. При взаимодействии СО2 со щелочью объем смеси уменьшается, создается вакуум и палец присасывается к отверстию.
РЕАКТИВЫ. Дрожжи; растворы с массовыми долями: сахарозы или глюкозы 5 %, гидроксида натрия 10 %, раствор Люголя.
КОНТРОЛЬНЫЕ ВОПРОСЫ.
1. Биологическое окисление и его роль.
2. Стадии окисления глюкозы в клетках, место их локализации.
3. Характеристика анаэробной стадии окисления глюкозы.
4. Роль анаэробной стадии окисления глюкозы для животных, человека, растений и микроорганизмов.
5. Характеристика гомоферментативного молочнокислого брожения и его роли.
6. Характеристика гетероферментативного молочнокислого брожения и его роли.
7. Характеристика спиртового брожения и его роли.
8. Характеристика уксуснокислого брожения и его роли.
9. Характеристика химизма пропионовокислого брожения и его роли.
10. Использование брожения в пищевой промышленности.
11. Роль брожения в живой природе.
Глава 5. Белки и их обмен
Гидролиз белков в организме идет постоянно как в процессе пищеварения (гидролиз белков пищевых продуктов), так и в процессе жизнедеятельности клеток (гидролиз устаревших или «изношенных» белков, а также запасных белков семян, зерен злаковых, клубней и других органов возрождения растений) протеазами. Конечными продуктами гидролиза белков являются аминокислоты.
megalektsii.ru
Аэробное окисление углеводов
Дыхание. Аэробное окисление углеводов происходит в присутствии кислорода воздуха, в связи с чем его часто называют дыханием.
В отличие от гликолиза (гликогенолиза), где конечным акцептором атомов водорода и электронов служит провиноградная кислота, при дыхании роль такого акцептора выполняет кислород. В первом случае в качестве конечного продукта образуется молочная кислота, в которой суммарная степень окисления углерода осталась такой же, как и у глюкозы, во втором случае образуется углекислый газ — значительно более простое соединение, у которого единственный атом углерода полностью окислен. Вместе с тем дыхание и гликолиз имеют много общих звеньев.
Дыхание, так же как и гликолиз, сопровождается образованием фосфорных эфиров глюкозы и фруктозы, фосфотриоз—диоксиацетонфосфата и глицеральдегид-3-фосфата, а также таких промежуточных продуктов, как 1,3-дифосфоглицериновая кислота, 3-фосфоглицерат, фосфоенолпирувати пировиноградная кислота. Многие реакции гликолиза и дыхания катализируются одними и теми же ферментами. Другими словами, при дыхании превращение глюкозы до молочной кислоты проходит все те этапы, что и при гликолизе. Однако при этом атомы водорода, отщепленные от глицеральдегид-3-фосфата, не восстанавливают пировиноградную кислоту, а передаются на кислород, пройдя через сложную систему ферментов дыхательной цепи.
Молочная кислота, образующаяся в процессе гликолиза, как уже говорилось, содержит еще довольно значительный запас (примерно 93 %) потенциальной энергии. Однако несмотря на это, первые живые организмы, извлекавшие энергию в анаэробных условиях, выделяли ее в окружающую среду.
С появлением в атмосфере Земли кислорода живые организмы выработали новые, более совершенные механизмы окисления, в результате которых количество высвобождающейся энергии оказалось намного больше, чем при гликолизе, поскольку конечным продуктом дыхания является СО2, атом углерода которого полностью окислен. Наряду с этим природа создала новые механизмы доокисления конечного продукта гликолиза, который выводился в окружающую среду. Иными словами, она как бы создала надстройку над гликолизом для окисления его конечного продукта в аэробных условиях, сохранив прежними многие его этапы.
При дыхании не образуется молочная кислота. Поэтому пировиноградная кислота является тем общим субстратом, или центральным звеном, где заканчивается гликолиз и начинается дыхание (или же расходятся пути гликолиза и дыхания — анаэробного и аэробного окисления глюкозы).
Сохранив прежние этапы гликолиза, клетки организма человека и высших животных сохранили способность окислять глюкозу в анаэробных условиях, в результате чего при недостатке кислорода они имеют возможность получать энергию таким путем. Однако при этом образовавшаяся в анаэробных условиях молочная кислота, обладающая довольно большим запасом энергии, не выбрасывается в окружающую среду, а накапливается и мышцах. Из мышц она током крови доставляется в печень, где снова превращается в глюкозу. При поступлении в клетку достаточного количества кислорода часть молочной кислоты окисляется дальше до СО2 и Н2О.
Превращение молочной кислоты. Образовавшаяся при анаэробном окислении глюкозы молочная кислота окисляется до СО2 и Н2О следующим образом. Сначала под действием фермента лактатдегидрогеназы, коферментом которой является НАД, она окисляется до пировиноградной кислоты:
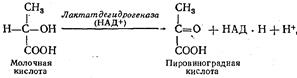
которая затем под влиянием пируватдекарбоксилазы, представляющей собой сложный полиферментный комплекс, подвергается окислительному декарбоксилированию с образованием активной формы уксусной кислоты — ацетил- КоА:
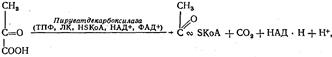
где ТПФ — тиаминпирофосфат; ЛК — липоевая кислота; HSKoA — коэнзим А.
В том случае, когда ткани хорошо снабжаются кислородом, пировиноградная кислота подвергается окислительному декарбоксилированию сразу, не восстанавливаясь до молочной кислоты. Восстановленный же кофермент НАД • Н + Н+, образовавшийся при окислении глицеральдегид-3-фосфата, передает водород через ферменты аэробного обмена (т.е. дыхательную цепь) на кислород, образуя воду.
Превращение пировиноградной кислоты в ацетил-КоА является подготовительной, или переходной, стадией, благодаря которой углеводы через пировиноградную кислоту, а затем через ацетил-КоА включаются в новый этап — кислородное окисление. Другими словами, этот процесс — связующее звено между гликолизом и собственно дыханием. Однако уже в результате окислительного декарбоксилирования пировиноградной кислоты до ацетил-КоА высвобождается около 9 % всей энергии окисления глюкозы, т.е. больше, чем при гликолизе в целом, где высвобождается всего 5—7 % энергии. Если учесть 5—7 % энергии гликолиза и 9 % энергии окислительного декарбоксилирования пировиноградной кислоты, то всего выделяется 14—16 % энергии, аккумулированной в углеводах. Следовательно, остальные 84—86 % энергии сохраняется еще в молекуле уксусной кислоты.
Цикл трикарбоновых кислот (цикл Кребса) представляет собой новый, более совершенный механизм окисления углеводов, выработанный у живых организмов с появлением на Земле кислорода. При помощи этого механизма происходит дальнейшее превращение уксусной кислоты в форме ацетил-КоА до СО2 и Н2О в аэробных условиях с высвобождением энергии.
В связи с тем что первыми субстратами в процессе окисления уксусной кислоты являются трикарбоновые кислоты, а гипотезу о механизме этого окисления выдвинул X. А. Кребс, процесс назвали циклом трикарбоновых кислот, или циклом Кребса.
Первой реакцией цикла является реакция конденсации ацетил-КоА со щавелевоуксусной кислотой, которую катализирует фермент цитратсинтаза. В результате образуется активная форма лимонной кислоты — цитрил-KoA:

Гидролизуясь, цитрил-КоА превращается в лимонную кислоту:
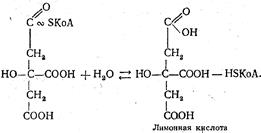
Последняя под действием фермента аконитатгидратаза превращается в цис-аконитовую кислоту, которая, присоединяя воду, превращается в изолимонную кислоту:

Изолимонная кислота далее окисляется путем отщепления двух атомов водорода, превращаясь в щавелевоянтарную. Этой реакцией начинается отщепление СО2 и первое окисление ацетил-КоА в трикарбоновом цикле. Щавелево-янтарная кислота, декарбоксилируясь, превращается в α-кетоглутаровую кислоту. Дегидрирование изолимон-ной и декарбоксилирование щавелево-янтарной кислот катализируется ферментом изоцитратдегидрогеназой с участием кофермента НАД+:

Следующим этапом цикла трикарбоновых кислот является реакция окислительного декарбоксилирования α-кетоглутаровой кислоты, в результате которой образуется янтарная кислота. Этот процесс протекает в две стадии. Сначала α-кетоглутаровая кислота подвергается окислительному декарбоксилированию с образованием активной формы янтарной кислоты — сукцинил-КоА — и СО2. Эта реакция напоминает реакцию превращения пировиноградной кислоты до ацетил-КоА и катализируется также сложным полиферментным комплексом — α-кетоглутаратдегидрогеназой. В результате этой реакции происходит второе отщепление углекислого газа и дегидрирование уксусной кислоты, вступившей в цикл:

Образовавшаяся активная форма янтарной кислоты сукцинил-КоА, в отличие от ацетил-КоА, представляет собой макроэргическое тио-эфирное соединение, в котором аккумулирована энергия окисления α-кетоглутаровой кислоты.
На следующей стадии эта энергия используется для образования ГТФ (гуанозинтрифосфорной кислоты) из ГДФ и неорганической фосфорной кислоты и запасается в фосфатных связях этого соединения. Реакция катализируется ферментом сукцинилтиокиназой:

Образовавшийся в результате этой реакции ГТФ взаимодействует с АДФ, в результате чего образуется АТФ:
ГТФ + АДФ  ГДФ + АТФ.
ГДФ + АТФ.
Синтез АТФ, сопряженный с окислением субстрата, является еще одним примером субстратного фосфорилирования.
В дальнейшем ходе цикла трикарбоновых кислот происходит еще два дегидрирования. Янтарная кислота под действием сукцинатде-гидрогеназы с участием кофермента ФАД+ отщепляет два атома водорода и превращается в фумаровую кислоту, а ФАД+ восстанавливается до ФАД•Н2. Затем фумаровая кислота, присоединяя молекулу воды, образует яблочную кислоту (малат), которая при помощи малат-дегидрогеназы и кофермента НАД+ снова подвергается дегидрированию. При этом образуется щавелево-уксусная кислота, т.е. субстрат, с которого начался цикл трикарбоновых кислот:

Регенерированная щавелево-уксусная кислота может снова вступать в реакцию с новой молекулой ацетил-КоА, и процесс начнется в том же порядке.
Общую схему цикла трикарбоновых кислот можно представить следующим образом:
Цикл трикарбоновых кислот

(в рамках показаны конечные продукты окисления ацетил-КоА).
Из приведенной схемы следует, что основная функция цикла Кребса заключается в дегидрировании уксусной кислоты. Если подвести баланс ферментативного дегидрирования одного цикла, можно легко подсчитать, что в результате реакций образуется восемь атомов водорода: шесть атомов используется для восстановления НАД+ и два — для восстановления ФАД+ сукцинатгидрогеназы.
Суммарная реакция этого цикла описывается следующим уравнением:
СН3СООН + 2Н2О →2СО2 + 8Н,
из которого следует, что четыре атома водорода принадлежат воде. Следовательно, остальные четыре образовались при дегидрировании уксусной кислоты, т.е. это весь водород, который был в составе ее молекулы. Одновременно с этим в виде оксида углерода (IV) дважды выделилось два атома углерода (один раз при декарбоксилировании щавелево-янтарной кислоты, второй — при декарбоксилировании α-кетоглутаровой кислоты), т.е. ровно столько, сколько их поступило в цикл в виде ацетальной группы.
Из приведенного выше уравнения также следует, что в цикл не вовлекаются ни кислород, ни АТФ, ни неорганическая фосфорная кислота. Все эти метаболиты взаимодействуют в дыхательной цепи, куда вовлекаются неорганическая фосфорная кислота, отщепленные при дегидрировании атомы водорода и кислород, а в результате окислительного фосфорилирования образуется АТФ. Энергия для этого процесса выделяется в результате окислительно-восстановительных реакций при передаче атомов водорода и электронов от восстановленных форм НАД • Н2 и ФАД • Н2 на кислород.
Процесс окислительного фосфорилирования подробно изложен в гл. 22. Напомним только, что на каждую пару электронов (пара атомов водорода) в дыхательной цепи путем окислительного фосфорилирования образуется три молекулы АТФ (одна при переносе атомов водорода от НАД • Н + Н+ к ФАД, вторая — при переносе пары электронов от цитохрома b к цитохрому с и третья — от цитохрома а3к атому кислорода). Таким образом, каждая окислительная стадия превращения глюкозы до СО2 и Н2О, связанная с НАД, сопровождается образованием трех молекул АТФ, связанная с ФАД — образованием двух молекул АТФ.
Энергетический баланс окисления углеводов.Сначала подведем итог энергетического баланса за счет дегидрирования уксусной кислоты в цикле Кребса. Как мы уже установили, в этом цикле происходит четыре дегидрирования, в результате которых образовались три восстановленные формы НАД, одна— ФАД и путем субстратного фосфорилирования синтезировалась одна молекула АТФ:
| 3 НАД • Н + Н+ | — 9 молекул АТФ |
| 1 ФАД • Н2 | — 2 молекулы АТФ |
| Фосфорилирование на уровне субстрата | — 1 молекула АТФ |
| Всего: | 12 молекул АТФ. |
Таким образом, в цикле Кребса синтезируется в шесть раз больше АТФ, чем при гликолизе. Если учесть еще две восстановленные молекулы НАД, образовавшиеся при окислении молочной и пировиноград-ной кислот, то это составит еще 6 молекул АТФ, а в сумме— 18. Поскольку глюкоза распадается на две фосфотриозы, количество АТФ увеличивается в 2 раза и составит 36 молекул.
Добавив к этому 2 молекулы АТФ, образовавшиеся в процессе гликолиза, получим общий баланс энергии, аккумулированной в мак-роэргических связях АТФ при окислении глюкозы до СО2 и Н2О: 36 + 2 = 38.
Установлено, что полное окисление 1 моль глюкозы до СО2 и Н2О сопровождается выделением 2872 кДж. В 38 молекулах АТФ аккумулируется 1270—1560 кДж, т.е. приблизительно 50 % всей энергии, высвободившейся в процессе окисления. Следовательно, остальные 50 % энергии рассеиваются в организме в форме теплоты для поддержания соответствующей температуры.
Из рассмотренных фаз окисления глюкозы исключительно важное значение имеет аэробная фаза. Если при анаэробном окислении, т.е. при образовании молочной кислоты, выделяется всего 197 кДж энергии, из которых 40 % аккумулируется в макроэргических связях двух молекул АТФ, то в аэробной фазе выделяется 2872 — 197 = = 2675 кДж, что составляет около 93 % всей энергии. Таким образом, основную массу энергии организм получает при дыхании.
Апотомический путьокисления глюкозы. Наряду с циклом Кребса во многих клетках существует и другой путь расщепления глюкозы, называемый апотомическим, или пентозофосфатным. Экспериментально установлено, что в аэробных условиях в эритроцитах, печени, почках глюкоза может окисляться до 6-монофосфоглюконовой кислоты, причем фруктозо-1,6-дифосфат в этом процессе не образуется. В результате такого окисления глюкозы образуется значительное количество пентоз. Этот путь был открыт советским биохимиком В. А. Энгельгардтом, а отдельные его этапы изучены О. Варбургом, Ф. Диккенсом, И. Д. Головацким и др. Пентозофосфатный путь не является главным путем окисления глюкозы. Основное его назначение состоит в том, чтобы снабжать клетки восстановленными формами НАДФ, необходимыми для биосинтеза жирных кислот, холестерина, пуриновых и пиримидиновых оснований, стероидов и др. Вторая функция этого пути заключается в том, что он поставляет пентозы, главным образом D-рибозу, для синтеза нуклеиновых кислот.
Пентозофосфатный путь расщепления глюкозы суммарно можно выразить следующим уравнением:
Глюкозо-6-монофосфат + 2 НАДФ+ → Рибозо-5-монофосфат + СО2 + 2 НАДФ·Н + Н+ + 2Н+.
Пентозы, не использованные для биосинтеза нуклеиновых кислот и нуклеотидов, расходуются на биосинтез других соединений и регенерацию глюкозы.
Биосинтез углеводов
Существует два основных способа биосинтеза углеводов из относительно несложных метаболитов. Один из них заключается в восстановлении углекислого газа до глюкозы. Этот процесс, характерный для зеленых растений и называемый фотосинтезом, осуществляется за счет энергии солнечных лучей при помощи хлорофилла согласно следующему уравнению:
СО2 + 2Н2О 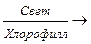 1/6С6Н12О6 + О2 + Н2О.
1/6С6Н12О6 + О2 + Н2О.
Улавливая солнечные лучи и преобразуя их энергию в энергию углеводов, зеленые растения обеспечивают сохранение и развитие жизни на Земле. В этом заключается, по словам К. А.Тимирязева, космическая роль зеленых растений как посредника между солнцем и всем живым на Земле.
В последнее время работами группы ученых Института биохимии им. А. В. Палладина АН УССР под руководством академика М. Ф. Гулого показано, что ткани высших животных также способны фиксировать углекислый газ, хотя механизм фиксации его отличается от такового у фотосинтезирующих клеток. Он заключается в наращивании углеродного скелета оксидом углерода (IV) таких субстратов, как кетокислоты, жирные кислоты, аминокислоты и др.
В печени, почках и скелетных мышцах человека и высших животных существует другой путь биосинтеза углеводов, называемый глюконеогенезом. Это синтез глюкозы из пировиноградной или молочной кислоты, а также из так называемых гликогенных аминокислот, жиров и других предшественников, которые в процессе метаболизма могут превращаться в пировиноградную кислоту или метаболиты цикла трикарбоновых кислот.
Глюконеогенез— это путь, обратный гликолизу. Однако в этом пути есть три стадии, которые в энергетическом отношении не могут быть использованы при превращении пировиноградной кислоты в глюкозу. Эти три стадии гликолиза заменены «обходными» реакциями с меньшей затратой энергии.
Первой обходной реакцией является превращение пировиноградной кислоты в фосфоенолпировиноградную. Поскольку расщепление глюкозы происходит в митохондриях, а синтез — в цитоплазме, на первом этапе митохондриальная пировиноградная кислота превращается сначала в щавелево-уксусную. Катализирует это превращение фермент пируваткарбоксилаза, активируемая ацетил-КоА с участием АТФ. Образовавшаяся щавелево-уксусная кислота восстанавливается затем с участием НАД • Н +Н+ в яблочную:
Пировиноградная кислота + СО2 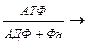 Щавелево-уксусная кислота
Щавелево-уксусная кислота  Яблочная кислота.
Яблочная кислота.
Яблочная кислота диффундирует в цитоплазму, окисляется цитоплазматической малатдегидрогеназой с образованием цитоплазматической щавелево-уксусной кислоты, из которой образуется фосфоенолпировиноградная кислота. Эту реакцию катализирует фосфоенолпируваткарбоксикиназа. Донором фосфорной кислоты служит ГТФ:
Яблочная кислота  Щавелево-уксусная кислота
Щавелево-уксусная кислота  Фосфоенолпировиноградная кислота.
Фосфоенолпировиноградная кислота.
Далее следует целая серия обратных реакций, заканчивающихся образованием фруктозо-1,6-дифосфата. Превращение фруктозо-1,6-дифосфата в фруктозо-6-фосфат— вторая необратимая реакция гликолиза. Поэтому она катализируется не фосфофруктокиназой, а фруктозодифосфатазой. Этот фермент катализирует необратимый гидролиз 1-фосфатной группы:
Фруктозо-1, 6-дифосфат + Н2О →Фруктозо-6-фосфат + Н3РО4.
На следующей (обратимой) стадии биосинтеза глюкозы фруктозо-6-фосфат превращается в глюкозо-6-фосфат под действием фосфоглюко-изомеразы гликолиза.
Расщепление глюкозо-6-фосфата до глюкозы — третья необратимая реакция, которая не осуществляется путем обращения гексокиназой. Свободная глюкоза образуется при помощи глюкозо-6-фосфатазы, катализирующей реакцию гидролиза:
Глюкозо-6-фосфат +Н2О → Глюкоза + h4PO4.
В большинстве клеток глюкозо-6-фосфат, образующийся в процессе гликогенолиза, используется как предшественник для биосинтеза олиго- и полисахаридов. Большую роль в биосинтезе этих сложных сахаров играет соединение уридинфосфоглюкоза, которая выполняет роль промежуточного переносчика глюкозы.
При биосинтезе гликогена, например, глюкозо-6-фосфат, превратившись в глюкозо-1-фосфат под действием фосфоглюкомутазы, взаимодействует с уридинтрифосфорной кислотой (УТФ) — соединением, аналогичным АТФ, в которое вместо аденина входит азотистое основание урацил. В результате этого взаимодействия при помощи глюкозо-I -фосфатуридилтрансферазы образуется уридилдифосфоглюкоза:
Глюкозо-1-фосфат + УТФ  УДФ-глюкоза+Фн.
УДФ-глюкоза+Фн.
На заключительном этапе биосинтеза гликогена в реакции, катали-зируемой гликогенсинтетазой, остаток глюкозы с УДФ-глюкозы переносится на концевой остаток глюкозы амилазной цепи с образованием 1,4-гликозидной связи (см. гл. 16). Ветвление гликогена путем образования 1,6-связей завершается амило-1,4-1,6-трансглюкозидазой.
Биосинтез гликогена осуществляется не только из глюкозо-6-фосфата, образовавшегося путем глюконеогенеза. Как уже отмечалось выше, для его биосинтеза используется также часть глюкозы после всасывания. Синтез гликогена, как процесс образования подвижного резерва углеводов в организме, имеет большое биологическое значение. Ведущая роль в этом принадлежит печени. Благодаря синтезу и отложению гликогена в печени поддерживается постоянная концентрация глюкозы в крови и других тканях, а также предотвращаются потери ее с мочой при употреблении пищи, особенно углеводной. Кроме того, отложение гликогена в печени способствует постепенному использованию углеводов в зависимости от условий существования организма.
Использованию глюкозы для синтеза гликогена предшествует образование глюкозофосфорных эфиров. Сначала образуется глюко-зо-6-монофосфат. Источником энергии и донатором фосфата является АТФ. Катализирует эту реакцию гексокиназа. Под действием фермента фосфоглюкомутазы глюкозо-6-монофосфат превращается в глюкозо-1-монофосфат:
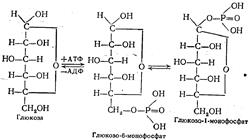
Дальнейшее превращение глюкозо-1-монофосфата до гликогена протекают уже знакомым нам путем.
Глава 24. ОБМЕН ЛИПИДОВ
Липиды представляют собой большую группу органических соединений. Все они различаются по своему химическому составу и структуре, но обладают одним общим для них свойством — нерастворимостью в воде. В связи с тем что ферменты, действующие на эти органические соединения, водорастворимы, расщепление и всасывание липидов в пищевом канале характеризуются некоторыми особенностями. Наличие же липидов различной структуры обусловливает различные пути их расщепления и синтеза.
Остановимся на обмене жиров, фосфатидов и стеридов, имеющих наиболее важное биологическое значение.
Обмен липидов, как и углеводов,— многоступенчатый процесс, который состоит из пищеварения, всасывания, транспортирования липидов кровью, внутриклеточного окисления и биосинтеза.
Переваривание липидов
Переваривание триглицеридов. Триглицериды, или нейтральные жиры, являются концентрированными источниками энергии в организме. При окислении 1 г жира высвобождается около 38,9 кДж энергии. Являясь гидрофобными соединениями, жиры резервируются в компактной форме, занимая сравнительно мало места в организме. Вместе с пищей в организм человека ежесуточно поступает до 70 г жиров растительного и животного происхождения. По своей химической природе они являются главным образом триглицеридами.
Расщепление жиров происходит при помощи ферментов, называемых липазами. Слюна не содержит таких ферментов, поэтому в ротовой полости жиры никаким изменениям не подвергаются. В желудке активность липазы очень слабая. Это связано с тем, что в желудке реакция среды сильнокислая (рН = 1,5-2,5), в то время как оптимум действия липазы находится при рН = 7,8 = 8,1. В связи с этим в желудке переваривается всего 3-5 % поступающих жиров.
Переваривание жиров в желудке происходит только у новорожденных и детей грудного возраста. Это связано с тем, что рН среды в желудке новорожденных составляет 5,6, а в этих условиях липаза проявляет большую активность. Кроме того, жир материнского молока, которое является основным продуктом питания детей в этот период, находится в сильно эмульгированном состоянии, а само молоко содержит липолитический фактор, принимающий участие в переваривании жиров.
Однако желудок все же играет определенную роль в процессе переваривания жиров у взрослых. Он регулирует поступление жира в кишки и переваривает белки, освобождая таким путем жир из липопротеидных комплексов пищи.
Основным местом переваривания жиров является двенадцатиперстная кишка и отделы тонкой кишки. Поскольку жиры нерастворимы в воде, а ферменты, расщепляющие их, являются водорастворимыми соединениями, необходимым условием для гидролитического расщепления жиров на составные части является их диспергирование (дробление) с образованием тонкой эмульсии. Диспергирование и эмульгирование жира происходит в результате действия нескольких факторов: желчных кислот, свободных высших жирных кислот, моно- и диглицеридов, а также белков. Этому способствуют также перистальтика кишок и постоянно образующийся углекислый газ, который выделяется при взаимодействии кислых компонентов пищи, поступающих из желудка, с карбонатами кишок, создающими щелочную среду. Образовавшийся углекислый газ «пробулькивает» через пищевые массы, участвуя таким образом в диспергировании жира. Нейтрализации содержимого желудка способствует также поступление в просвет тонкой кишки желчи, обладающей щелочным характером.)
Желчь — вязкая жидкость светло-желтого цвета со специфическим запахом, горькая на вкус. В состав желчи входят желчные кислоты. желчные пигменты, продукты распада гемоглобина, холестерин, лецитин, жиры, некоторые ферменты, гормоны и др. Желчь способствует перистальтике тонкой кишки, оказывает бактериостатическоедействие на ее микрофлору. С желчью выделяются из организма яды. Она является также активатором липолитических ферментов и повышает проницаемость стенки кишок.
Главной составной частью желчи являются желчные кислоты. Они образуются в печени из холестерина и находятся в желчи как в свободном, так и в связанном состоянии, а также в виде натриевых солей. В желчи человека содержится в основном три желчных кислоты Основную массу составляют холевая (3,7,12-тригидроксихола-новая) и дезоксихолевая (3,12-дигидроксихолановая), небольшую часть — литохолевая (3-гидроксихолановая) кислоты, которые являются производными холановой кислоты:

Холевая кислота может находиться в желчи также в связанном состоянии в виде парных соединений с глицином и производным цистеина таурином — соответственно гликохолевой и таурохолевой кислот:
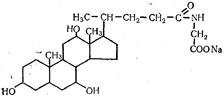 Натриевая соль гликохолевой кислоты
Натриевая соль гликохолевой кислоты
 Натриевая соль таурохолевой кислоты
Натриевая соль таурохолевой кислоты
Благодаря наличию желчных кислот происходит снижение поверхностного натяжения липидных капель, что способствует образованию очень тонкой и устойчивой эмульсии диаметр частиц которой составляет около 0,5 мкм. Образованию эмульсии способствуют также моноглицериды и высшие жирные кислоты. Эмульгирование жира приводит к колоссальному увеличению поверхности соприкосновения липазы с водным раствором. Таким образом, чем тоньше эмульсия жиров, тем лучше и быстрее они расщепляются липазой. Кроме того, в виде тонкой эмульсии жиры могут даже всасываться стенкой кишок непосредственно, не расщепляясь на составные части.
В присутствии желчных кислот под действием липазы в просвете тонкой кишки происходит гидролитическое расщепление жиров. В результате этого образуются продукты частичного и полного расщепления жиров — моно- и диглицериды, свободные высшие жирные кислоты и глицерин:
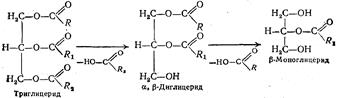

Здесь же содержится и часть нерасщепленного жира в виде очень тонкой эмульсии. Все эти продукты в дальнейшем всасываются стенкой кишок. В этой смеси триглицериды составляют около 10 % , моно-
идисахариды — также 10 % , а основная масса — около 80 % — продукты полного расщепления жиров— глицерин и высшие жирные
кислоты.
Переваривание фосфоглицеридов.Основным местом переваривания фосфатидов также является двенадцатиперстная кишка. Эмульгирование этих липидов происходит под влиянием тех же веществ, что и три-глицеридов. Однако гидролитическое расщепление фосфатидов осуществляется под действием фосфолипаз А, В, С и D. Каждый фермент действует на определенную сложноэфирную связь фосфолипида. Гидролитическое расщепление, например, лецитина происходит следующим образом:



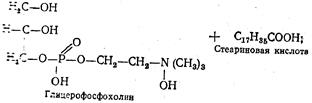
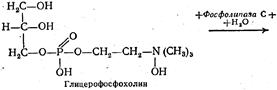



Такому полному расщеплению подвергается незначительная часть фосфатидов, поскольку его промежуточные продукты хорошо растворимы в воде и легко всасываются стенкой кишок. К тому же фосфогли-цериды легко образуют эмульсии, которые также могут всасываться кишечной стенкой.
Переваривание стеридов.Стериды, входящие в состав пищи, эмульгируются под влиянием тех же факторов, что и жиры, после чего подвергаются гидролитическому расщеплению до свободных стеринов и высших жирных кислот. Этот процесс осуществляется под действием фермента холестеринэстеразы.
Всасывание липидов
Врезультате пищеварения жиров, фосфатидов, стеридов в просвете тонкой кишки образуется значительное количество продуктов их частичного и полного гидролитического расщепления: моно- и диглицериды, высшие жирные кислоты, стерины, азотистые основания, фосфорная кислота. Содержится также небольшое количество триглицеридов, находящихся в тонкоэмульгированном состоянии. Все эти продукты всасываются стенкой тонкой кишки.
Такие продукты расщепления, как жирные кислоты и холестерин, плохо растворяясь в воде, образуют с желчными кислотами водорастворимые комплексы— так называемые холеиновые кислоты. Эти кислоты легко проникают в эпителиальные клетки стенки кишок, где расщепляются на составные части. Освобожденные желчные кислоты возвращаются в просвет кишок и снова используются для транспортирования нерастворимых в воде продуктов расщепления жиров.
Часть продуктов расщепления (глицерин, глицеринфосфорная кислота, азотистые основания) хорошо растворимы в воде и легко проникают в эпителиальные клетки. Фосфорная кислота всасывается в клетки эпителия стенки тонкой кишки в виде натриевых и калиевых солей. В основе всасывания липидов лежит ряд сложных физико-химических и биологических процессов, для осуществления которых затрачивается энергия макроэргических связей АТФ.
В эпителиальных клетках слизистой оболочки кишок из всосавшихся продуктов гидролитического расщепления снова синтезируются липиды. Однако этот ресинтез приводит к образованию специфических жиров, характерных для данного организма.
Для образования нейтральных жиров используются высшие жирные кислоты, глицерин, моно- и диглицериды. Одновременно происходит и синтез фосфатидов, для которых используются главным образом глицеринфосфорная кислота, глицериды и диглицериды, а также в небольшом количестве моноглицериды. Из холестерина и высших жирных кислот образуются стериды.
В эпителиальных клетках стенки кишок из синтезированных липидов, а также капель всосавшихся триглицеридов, витаминов (A, D, Е, К) ибелков образуются комплексы размером 150—200 нм, называемые хило микронами. Внутреннее содержимое хиломикрона, представленное образовавшимися различного рода липидами, главным образом триглицеридами, окружено наружной белковой оболочкой, благодаря которой хиломикроны хорошо растворяются в воде. Хило-микроны диффундируют сначала в межклеточную жидкость, затем в лимфатические капилляры и в конце концов попадают в кровяное русло, где под действием гепарина распадаются на мелкие частицы. С током крови они разносятся по всему организму и откладываются в резерв в жировых депо — подкожной и околопочечной клетчатке, сальнике, брыжейке, мышечной ткани. Часть жиров крови используется для пластических целей, как источник химической энергии и т.д.
Таким образом, хиломикроны являются переносчиками образовавшихся в эпителиальных клетках тонкой кишки липидов. При этом они транспортируют в крови главным образом триглицериды.
Наряду с хиломикронами существуют и другие формы транспорта липидов кровью, например α- и β-липопротеиды. Их молекулы представляют собой сложные комплексы липидов с белками. α-Липопро-теиды являются основными транспортными формами фосфатидов, β-липопротеиды— переносчиками холестерина и его эфиров.
Наиболее подвижной формой липидов являются свободные высшие жирные кислоты.
Важная роль в активном транспортировании липидов принадлежит форменным элементам крови. Эритроциты, например, участвуют в переносе фосфатидов и холестерина, лейкоциты— триглицеридов.
Большая роль в обмене липидов принадлежит жировым депо. Исследования показали, что в жировых депо откладывается не только вновь синтезированный в организме специфически видовой жир, но и в небольших количествах чужеродный, т.е. входящий в состав пищи. Опыты, проведенные на голодающих собаках, показали, что пищевые жиры после всасывания поступают сначала в жировые депо, из которых переходят в плазму крови.
Таким образом, жировая ткань не является пассивным депо жиров, состав ее постоянно обновляется за счет липидов, всасывающихся из кишок или синтезируемых в организме.
lektsia.com
Глюкоза Этиловый
спирт
2. Молочнокислое брожение идет по уравнению:
СбН12Об 2 СН3СНОН—СООН
3. Маслянокислое брожение:
СбН120б — СН3—СН2—СН2—СООН+2СО2+2Н2
Масляная кислота
Первые этапы при различных видах брожения идут аналогично вышеописанному анаэробному распаду глюкозы при гликолизе до образования пировиноградной кислоты. Так же, как это было рассмотрено выше для животных тканей, в микроорганизмах (дрожжах, бактериях) происходит прежде всего фосфорилирование глюкозы с образованием глюкозо-6-фосфат, а затем фруктозо-1,6-дифосфат. Дифосфат фруктозы также распадается на две молекулы глицеринового альдегида, а последний через ряд стадий дает пировиноградную кислоту.
Анаэробный распад углеводов в дрожжах (спиртовое брожение) приводит к образованию этилового спирта. При этом брожении пировиноградная кислота на первом этапе подвергается де-карбоксилированию под действием пируватдекарбоксилазы, кофер-ментом которой является тиаминдифосфат (тиаминпирофосфат). В результате образуется СО2 и уксусный альдегид, последний затем при участии восстановленной алкогольдегидрогеназы восстанавливается в спирт.
1. СН3—СО—СООН С02+СН3—С==0
Пировиноградная пируватдекарбок-
кислота силаза H
Уксусный
альдегид
2. СНз-С==O НАДН+Н+
алкогольдегидрогеназа С2Н5ОН + НАД+
Н
На процессе спиртового брожения основано виноделие и спирто-водочное производство.
Молочнокислое брожение идет при участии молочнокислых бактерий. Восстановленная дегидрогеназа (лактатдегидоогеназа) передает свой водород пировиноградной кислоте, которая восстанавливается в молочную кислоту.
лактатдегидро-
СН3-СО-СООН + 2Н СН3СH- ОН – СООН
геназа Молочная кислота
Молочнокислое брожение играет большую роль при производстве молочнокислых продуктов (кефира, простокваши и др.), при квашении капусты, огурцов, при силосовании кормов. Маслянокислое брожение вызывается маслянокислыми бактериями, большинство из которых являются анаэробами. Образование масляной кислоты также идет через пировиноградную кислоту.
Аэробное окисление углеводов.
Распад углеводов в аэробных условиях может идти прямым (aпотомическим или пентозным) путем и непрямым (дихотомическим) путем.
Дихотомическое (греч. dicha - на две части, tome-сечение) окисление углеводов идет по уравнению:
C6h22O6+6O2 6 СО2+б Н2О+686 ккал
Этот путь является основным в образовании энергии. Первые этапы этого пути совпадают с анаэробным окислением глюкозы. Расхождение путей начинается на стадии образования пировиноградной кислоты, которая в животных тканях декарбоксилируется окислительным путем.
Первоначально предполагали, что пировиноградная кислота подвергается окислительному декарбоксилированию с образованием уксусной кислоты по уравнению: СН3—СО—СООН + 1/2 O2 СН3СООН+СО2. Потом было установлено, что уксусная кислота не является промежуточным продуктом при .декарбоксилировании пировиноградной кислоты, и возникло представление, что уксусная кислота существует в «активной» форме. Вскоре было показано, что для утилизации пирувата необходим коэнизм А (КоА). В химическом отношении КоА представляет нуклеотид, в состав которого входит аденозин-31, 51-дифосфат, фосфат, пантотеновая кислота и тиоэтиламин. Коэнзим А участвует в переносе остатка уксусной кислоты — ацетильного радикала (Ch4CO-) также и других кислотных (ацильных) радикалов.
Окислительное декарбоксилирование пирувата — процеcc многоступенчатый, осуществляется сложной ферментативной системой, в состав которой помимо пируватдекарбоксилазы, тиаминдифосфата и коэнзима А входят дегидрогеназы с коферментом НАД+, и ФАД, липоевая кислота и ионы магния. В результатеокисления пировиноградной кислоты образуются молекула aцетил-КоА («активной» формы уксусной кислоты), два атома водорода (в виде НАДН+Н+ ) и молекула СО2.
O
СН3-СО-СООН+НS-КоА+НАД+СН3--С~S-~КоА+С02+НАДН+ H
Ацетил-КоА
Следующий этап непрямого аэробного окисления глюкозы характеризуется полным окислением ацетил-КоА в цикле Кребса до СО2 и Н2О.
ЦИКЛ ДИ- и ТРИКАРБОНОВЫХ КИСЛОТ (КРЕБСА).
Исследования показали, что дальнейшее окисление ацетил-КоА возможно лишь в присутствии небольших количеств какой-либо ди-карбоновой кислоты. Оказалось, что в начале ацетил-коэнзима А конденсируется с щавелевоуксусной кислотой (СООН—СН2— СО—СООН) с образованием лимонной (трикарбоновой) кислоты. Лимонная кислота является первым продуктом цикла Кребса, поэтому этот цикл иногда называют лимоннокислым.
Образовавшаяся лимонная кислота подвергается далее ряду сложных превращений. И прежде всего, дегидратированию с образованием цис-аконитовой кислоты. Последняя присоединяет молекулу воды и переходит в изолимонную кислоту. Изолимонная кислота подвергается дегидрированию и превращается в щавелево-янтарную, которая декарбоксилируется с образованием -кетоглютаровой.-Кетоглютаровая кислота подвергается окислительному декарбоксилированию и одновременно дегидрируется, переходя в сукцинил-КоА. Затем сукцинил-КоА превращается в янтарную кислоту. Янтарная кислота дегидрируется, превращаясь в фумаровую. Фумаровая переходит в яблочную, а из яблочной при ее дегидрировании образуется щавелевоуксусная. На этом цикл замыкается. Многие реакции цикла Кребса легко обратимы. Ферменты, катализирующие реакции цикла Кребса сосредоточены в митохондриях. Последовательность реакций цикла Кребса последовательно изображена на суммарной схеме (рис. 7.3) .
Из приведенной схемы видно, что в цикле Кребса в результате реакций дегидрирования образуется 4 пары водородных атомов и 2 молекулы СО2. Освободившийся в ходе процесса окисления водород поступает в цепь биологического окисления и в конечном итоге окисляется молекулярным кислородом с образованием воды и выделением энергии.
При окислении в цикле ди- и трикарбоновых кислот одной молекулы ацетил-КоА образуется 12 молекул АТФ, из которых одиннадцать возникает путем окислительного фосфорилирования, а одна при субстратном фосфорилировании (при превращении сукцинил-КоА в янтарную кислоту).При окислении большинства субстратов в цепи биологического окисления происходит образование 3-х молекул АТФ, тогда как окисление некоторых из них (например, в случае янтарной кислоты) дает 2 молекулы АТФ. Энергетический баланс анаэробного и аэробного окисления глюкозы представляет следующую картину.
1. Две молекулы АТФ — это чистый прирост АТФ при превращениях глюкозы до пировиноградной кислоты в анаэробной фазе.
Глюкоза+2 НАД+ +2 АДФ+2 Фн- 2 Пируват+2 НАДН+2 АТФ
2. Четыре молекулы АТФ образуются в результате окисления двух молекул НАДН, возникших при дегидрировании двух молекул 3-фосфоглицеринового альдегида (рис. 13). В связи с тем, что эти две молекулы НАДН являются цитоплазматическими, то отдаваемые ими электроны могут включиться в митохондриальную цепь биологического окисления не прямым путем, а с помощью так называемого челночного механизма. Суть этого механизма состоит в том, что сначала цитоплазматический НАДН реагирует с фосфодиоксиацетоном и образует глицерол-3-фосфат.
Фосфодиоксиацетон+НАДН Глицерол-3-фосфат+НАД+
Г'лицерол-3-фосфат легко проникает через митохондриальную мембрану и окисляется с участием флавинзависимой дегидрогеназы в фосфодиоксиацетон, причем простетическая флавиновая группа восстанавливается.
Глицерол-3-фосфат+Фл. пр. Фосфодиоксиацетон+
4-фл. пр. Н2. Фосфодиоксиацетон выходит из митохондрий, а восстановленный флавопротеид (Фл. пр. Н2) передает приобретенные электроны в цепь биологического окисления, обеспечивая окислительное фосфорилирование только двух молекул АДФ.
3. Шесть молекул АТФ возникают в процессе окислительного де-карбоксилирования двух молекул пировиноградной кислоты, образовавшихся в анаэробной .фазе из одной молекулы глюкозы.
4. При полном окислении двух молекул ацетил-КоА в цикле Кребса возникает 24 молекулы АТФ. В итоге полного аэробного окисления одной молекулы глюкозы синтезируется 36 молекул ЛТФ. При анаэробном гликолизе (брожении) на одну молекулу глюкозы образуется всего 2 молекулы АТФ. Таким образом, «выход» энергии, запасаемой в виде АТФ при кислородном распаде глюкозы, в 18 раз больше, чем при анаэробном.
ПЕНТОЗО-ФОСФАТНЫЙ ПУТЬ ОКИСЛЕНИЯ УГЛЕВОДОВ.
Существует еще один путь аэробного окисления углеводов, который имеет значение лишь в некоторых органах и тканях (жировой ткани, печени, эритроцитах, молочной железе, коре надпочечников, половых железах). Этот путь называется пентозофосфатным (пентозным, прямым или гексозомонофосфатным).
В отличие от непрямого (гексозодифосфатного) пути окисления углеводов в пентозном цикле не проходит стадии образования фруктозе-16-дифосфата, фосфотриоз и т. д., т. е. не происходит фосфорилирования глюкозомонофосфата. Последний в пентозном цикле подвергается прямому окислению с образованием в конечном итоге СО2 и пентозофосфата.
Значение пентозного цикла состоит, во-первых, в том, что является основным поставщиком восстановленного НАДФ + (НАДФН), необходимого для разнообразных синтетических процессов. Во-вторых, этот цикл обеспечивает организм пентозами. В третьих, энергетическая ценность цикла весьма велика — равна 36 мол АТФ.
7.5. ФОТОСИНТЕЗ
Фотосинтез - это синтез органических веществ (прежде всего углеводов) из углекислого газа и воды, происходящий за счет энергии света. Фотосинтез является основным источником образования органических веществ на Земле и единственным источником кислорода.
Процесс фотосинтеза в растениях можно выразить следующим
свет
суммарным уравнением: 6С02+6Н20 С6 Н 12О6+6 O2
хлорофилл
фотосинтез происходит внутри специализированных органелл- пластидах (хлоропластах), которые содержат в себе хлорофилл.
Хлорофилл — сложное гетероциклическое соединение, содержащее четыре перрольных кольца, образующих порфириновое ядро. Перрольные кольца связаны двумя основными и двумя дополнительными валентностями с атомом магния. Хлорофилл благодаря наличию в его структуре сопряжённых двойных связей способен поглощать световую энергию, переходя в активное состояние. Аккумулирование солнечной энергии хлорофиллом является начальным этапом фотосинтеза, который можно изобразить следующим образом:
Хл + hv — Хл+
Хлоро- Квант Хлорофилл
филл света возбужденный
На втором этапе фотосинтеза возбужденный хлорофилл, обогащенный энергией за счет поглощения светового фотона, отдаёт энергию на разложение воды (фотолиз воды). В результате образуются активные радикалы Н и ОН. Радикалы ОН дают перекись (ОН)2, распад которой приводит к образованию кислорода.
Хл+ +Н20 — Хл+Н+ОН
ОН (ОН)2 —— h3O+1/2O2
Следующий (третий) этап состоит в переносе водорода воды
через цепь переносчиков на НАДФ с образованием НАДФН. Одновременно происходит за счет переноса электронов, индуцируемого светом, синтез АТФ из АДФ и неорганического фосфата, т. е так называемое фотосинтетическое фосфорилирование.
Разобранные выше этапы являются световой фазой фотосинтеза. В реакциях этой фазы принимает участие поглощенная хлорофиллом световая энергия. Световая фаза приводит к образованию молекулярного кислорода, НАДФН и АТФ.
Следующая фаза фотосинтеза — темновая. Реакции в этой фазе происходят без участия света. В темновой фазе фотосинтеза НАДФН и АТФ используются для восстановления углекислого газа до углевода.
studfiles.net
Обмен и функции углеводов. Анаэробное и аэробное окисление глюкозы
15.2.1. Гликолиз – это ферментативный распад глюкозы в аэробных условиях до двух молекул пировиноградной кислоты (аэробный гликолиз), а в анаэробных условиях – до двух молекул молочной кислоты (анаэробный гликолиз). В анаэробных условиях гликолиз протекает в тканях без потребления кислорода и является единственным процессом, поставляющим АТФ, так как окислительное фосфорилирование в этих условиях не функционирует. Анаэробный гликолиз происходит во всех тканях, функционирующих в условиях гипоксии, прежде всего в скелетных мышцах. Гликолиз в эритроцитах даже в присутствии кислорода завершается образованием лактата, поскольку в этих клетках отсутствуют митохондрии.
Гликолиз протекает в цитозоле клеток организма. Этот процесс катализируется одиннадцатью ферментами, которые выделены в высокоочищенном состоянии и хорошо изучены. Условно можно разделить гликолиз на две стадии.
15.2.2. Первая стадия гликолиза является подготовительной и включает реакции превращения молекулы глюкозы в две молекулы фосфотриоз. Эта стадия сопровождается затратой молекул АТФ.
Начальной реакцией превращения глюкозы в клетке является её фосфорилирование в результате взаимодействия с АТФ (рисунок 15.1, реакция 1). Эта реакция в условиях клетки протекает только в одном направлении. Биологическая роль реакции фосфорилирования глюкозы заключается в том, что глюкозо-6-фосфат, в отличие от свободной глюкозы, не может проникать через плазматическую мембрану обратно в кровь и оказывается «запертой» в клетке. Таким образом, глюкозо-6-фосфат является ключевым метаболитом углеводного обмена, на уровне которого осуществляется интеграция различных путей превращения глюкозы в клетке.
В большинстве тканей реакцию фосфорилирования глюкозы катализирует фермент гексокиназа, которая обладает высоким сродством к глюкозе, способна также фосфорилировать фруктозу и маннозу и аллостерически ингибируется избытком глюкозо-6-фосфата. В клетках печени, кроме того, есть фермент глюкокиназа, которая имеет низкое сродство к глюкозе, не ингибируется глюкозо-6-фосфатом и не участвует в фосфорилировании других моносахаридов. Глюкокиназа эффективно функционирует только при высокой концентрации глюкозы в крови. Это способствует усвоению большого количества углеводов, поступающих в печень из кишечника в активную фазу пищеварения.
В следующей реакции глюкозо-6-фосфат изомеризуется во фруктозо-6-фосфат (рисунок 15.1, реакция 2).
Продукт реакции изомеризации подвергается повторному фосфорилированию за счёт АТФ (рисунок 15.1, реакция 3). Эта реакция – наиболее медленно протекающая реакция гликолиза и, подобно фосфорилированию глюкозы, необратима. Фермент – фосфофруктокиназа – является аллостерическим, активируется АДФ, АМФ, и фруктозо-2,6-бисфосфатом, а ингибируется цитратом и высокой концентрацией АТФ.
На следующем этапе фруктозо-1,6-дифосфат подвергается расщеплению на две фосфотриозы (рисунок 15.1, реакция 4). Таким образом, химическое соединение, содержащее 6 углеродных атомов, превращается в два, содержащих по 3 атома углерода. Поэтому гликолиз называют дихотомическим путём превращения глюкозы (от слова «дихотомия» – рассечение на две части).
Далее происходит изомеризация триозофосфатов (рисунок 15.1, реакция 5). В этой реакции диоксиацетонфосфат переходит в глицеральдегид-3-фосфат. Таким образом, в первой стадии гликолиза молекула глюкозы превращается в две молекулы глицеральдегид-3-фосфата. Поэтому в реакциях второй стадии глюкозы будет участвовать по две молекулы каждого субстрата, что необходимо учитывать при расчёте энергетического баланса данного метаболического пути.
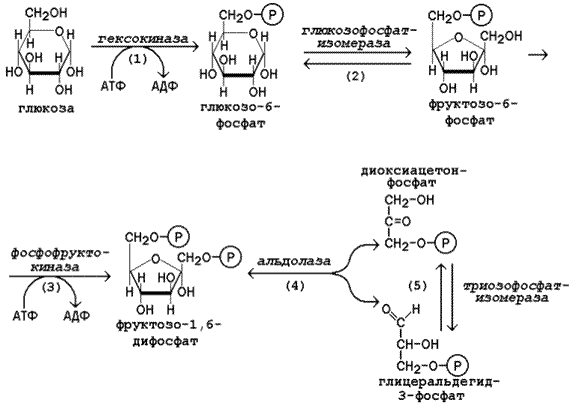
Рисунок 15.1. Реакции первой стадии гликолиза.
15.2.3. Вторая стадия гликолиза включает реакции превращения двух молекул глицеральдегид-3-фосфата в две молекулы лактата. На этой стадии гликолиза происходит синтез молекул АТФ.
Глицеральдегид-3-фосфат подвергается дегидрированию при участии НАД-зависимой дегидрогеназы. В этой реакции происходит потребление неорганического фосфата, который включается в состав продукта реакции, содержащего макроэргическую фосфатную связь (рисунок 15.2, реакция 6), а промежуточным донором водорода служит SH-группа в активном центре фермента, которая потом регенерирует.
1,3-Дифосфоглицерат вступает в реакцию первого субстратного фосфорилирования, т.е. не сопряжённого с переносом электронов в дыхательной цепи. В этой реакции осуществляется синтез молекулы АТФ в результате переноса фосфатной группы вместе с макроэргической связью на молекулу АДФ (рисунок 15.2, реакция 7).
В следующей реакции происходит внутримолекулярное перемещение фосфатной группы 3-фосфоглицерата ко 2-му углеродному атому (рисунок 15.2, реакция 8). Тем самым облегчается последующее отщепление молекулы воды, которое приводит к появлению в продукте реакции макроэргической фосфатной связи (рисунок 15.2, реакция 9).
Фосфоенолпируват (ФЕП) вступает в реакцию второго субстратного фосфорилирования, в ходе которого образуется молекула АТФ. В отличие от первого субстратного фосфорилирования, данная реакция является необратимой в условиях клетки (рисунок 15.2, реакция 10). Фермент пируваткиназа существует в двух изоферментных формах. Изофермент, присутствующий в печёночных клетках, аллостерически ингибируется АТФ и активируется фруктозо-1,6-дифосфатом. Изофермент, присутствующий в головном мозге, мышцах и других тканях, не является аллостерическим и не принимает участия в регуляции гликолиза.
В заключительной реакции гликолиза происходит использование НАДН, образовавшегося при дегидрировании глицеральдегид-3-фосфата (см. реакцию 6). При участии НАД-зависимой лактатдегидрогеназы пируват восстанавливается в молочную кислоту (рисунок 15.2, реакция 11). Фермент существует в пяти изоферментных формах, отличающихся сродством к субстрату и распределением в тканях.
 Рисунок 15.2. Реакции второй стадии гликолиза.
Рисунок 15.2. Реакции второй стадии гликолиза.
Таким образом, в процессе образования лактата из глюкозы в клетке не накапливается НАДН. Это значит, что данный процесс является анаэробным и может протекать без участия кислорода (который является конечным акцептором электронов, передаваемых НАДН в дыхательную цепь). В тканях, функционирующих в условиях гипоксии,
При подсчёте энергетического баланса гликолиза следует учитывать, что каждая из реакций второй стадии этого метаболического пути повторяется дважды. Таким образом, в первой стадии было затрачено 2 молекулы АТФ, а во второй стадии путём субстратного фосфорилирования образовалось 2х2 = 4 молекулы АТФ; следовательно при окислении одной молекулы глюкозы в клетке накапливается 2 молекулы АТФ.
dendrit.ru
Анаэробное окисление углеводов
Анаэробное окисление углеводов происходит в клетках, органах и тканях без участия кислорода. Если процесс начинается с превращения глюкозы и заканчивается образованием молочной кислоты, то он называется анаэробным гликолизом, если начинается с превращения гликогена – гликогенолизом.
Гликолиз
На 1-й стадии гликолиза (рис. 7) происходит фосфорилирование глюкозы под действием фермента гексокиназы в присутствии АТФ и ионов Mg2+ с образованием глюкозо-6-фосфата (Г-6-Ф), который на 2-й стадии превращается во фруктозо-6-фосфат (Ф-6-Ф). Эту реакцию катализирует глюкозофосфатизомераза. В ходе 3-й стадии осуществляется фосфорилирование фруктозо-6-фосфата, в результате чего образуется фруктозо-1,6-дифосфат (Ф-1,6-Ф). Катализ данной реакции обеспечивает в присутствии ионов Mg2+ и АТФ фосфофруктокиназа.
На 4-й стадии Ф-1,6-Ф под действием альдолазы расщепляется на 2 фосфотриозы - глицеральдегид-3-фосфат (ГА-3-Ф) и дигидроксиацетонфосфат (ДАФ).
ДАФ под действием триозофосфатизомеразы превращается в ГА-3-Ф. Таким образом, из одной молекулы глюкозы мы получаем 2 молекулы ГА-3-Ф.
ГА-3-Ф в присутствии глицеральдегидфосфатдегидрогеназы, кофермента НАД+ и фосфорной кислоты окисляется с образованием 1,3-дифосфоглицерата (1,3-ДФГ) и НАДН(Н+). Энергия, высвобождающаяся в этой реакции аккумулируется в макроэргической связи 1,3-ДФГ, который далее превращается в 3-фосфоглицерат под действием фосфоглицераткиназы. Этот процесс сопряжен с фосфорилированием АДФ (образуется АТФ на уровне субстратного фосфорилирования).
3-ФГ при участии фосфоглицератмутазы превращается в 2-фосфоглицерат (2-ФГ), который подвергается дегидратации в присутствии енолазы и ионов Mg2+. В результате образуется фосфоенолпируват (ФЕП), содержащий макроэргическую связь. ФЕП под действием пируваткиназы при наличии ионов Mg2+ превращается в енольную форму пирувата. Этот процесс сопряжен с синтезом АТФ на уровне субстратного фосфорилирования. Енольная форма пирувата таутомеризуется в кетонную форму.
На заключительной стадии гликолиза происходит восстановление пирувата до L-лактата в присутствии НАДН(Н+) и фермента лактатдегидрогеназы. Образующийся в этой реакции кофермент НАД+ в дальнейшем используется в реакции окисления ГА-3-Ф.
Таким образом конечными продуктами гликолиза являются 2 молекулы L-лактата и 2 молекулы АТФ в расчете на 1 молекулу глюкозы.
Регуляция гликолиза осуществляется на уровне фосфофруктокиназной реакции. Фермент ингибируется высоким содержанием АТФ и цитрата. Аллостерическими активаторами фосфофруктокиназы являются АМФ, АДФ, Ф-6-Ф.
Гликогенолиз
Гликоген представляет собой разветвленный полисахарид, состоящий из остатков -D-глюкозы, связанных между собой в линейных участках молекулы -1,4-гликозидными связями, а в точках ветвления -1,6-гликозидными связями.
Под действием гликогенфосфорилазы происходит отщепление одного глюкозного остатка в линейных участках с переносом его на молекулу фосфорной кислоты, в результате чего образуется глюкозо-1-фосфат (Г-1-Ф). Гликогенфосфорилаза работает до тех пор, пока до ближайшей точки ветвления не останется 4 остатка глюкозы, затем в работу включается фермент олигосахаридтрансфераза, переносящий фрагмент из 3-х глюкозных остатков на соседнюю ветвь, таким образом, в точке ветвления остается один глюкозный остаток. Он отщепляется гидролитическим путем с помощью -1,6-гликозидазы в виде свободной молекулы глюкозы и вновь создаются условия для работы гликогенфосфорилазы в линейной цепи.
Продукт глигогенфосфорилазной реакции Г-1-Ф далее под действием фосфоглюкомутазы превращается в Г-6-Ф, который вовлекается в гликолитический путь (рис. 8).
Рис. 8. Схема распада гликогена в мышцах и печени.
Конечные продукты гликогенолиза – 2 молекулы L-лактата и 3 молекулы АТФ в расчете на 1 молекулу глюкозы.
studfiles.net
Окислительные пути распада углеводов в тканях
Важнейшими функциями моносахаридов в организме являются энер-
гетическая и пластическая; обе эти функции реализуются в ходе
окислительного распада моносахаридов в клетках. При окислении углеводов выделяется 4,1 ккал/г ( около 17 кДж/г ) свободной энергии и за счет окисления углеводов человек покрывает 55-60% своих общих энергозатрат. В ходе окисления углеводов образуется большое количество промежуточных продуктов распада, которые используются для синтеза различных липидов, заменимых аминокислот и др. необходимых клеткам соединений. Кроме того, при окислении углеводов в клетках идет генерация восстановительных потенциалов, которые в дальнейшем используются ими в восстановительных реакциях биосинтезов, в процессах детоксикации, для контроля уровня перекисного окисления липидов и др.
Главным моносахаридом, подвергающимся окислительным превращениям в клетках, является глюкоза, поскольку именно она в наибольших количествах поступает из кишечника во внутреннюю среду организма, именно она синтезируется при глюконеогенезе или образуется в свободном виде или же в виде фосфорных эфиров при расщеплении гликогена. Роль других моносахаридов менее значительна, так как их количество, поступающее в клетки в количественном отношении сильно варьирует в зависимости от состава пищи.
Известно несколько метаболических путей окисления глюкозы, главными из которых являются:
а) аэробное расщепление до углекислого газа и воды;
б) анаэробное окисление до лактата;
в) пентозный путь окисления;
г) окисление с образованием глюкуроновой кислоты.
Глубина окислительного расщепления молекулы глюкозы может быть различной: от окисления одной из концевых группировок молекул до карбоксильной группы, что происходит при образовании глюкуроновой кислоты, до полной деградации молекулы глюкозы при ее аэробном распаде.
Аэробное окисление глюкозы
В клетках аэробных организмов основным, по крайней мере в отношении общего количества расщепляющейся глюкозы, является ее аэробный распад до углекислого газа и воды. При расщеплении 1 М глюкозы ( 180 г ) в аэробных условиях выделяется 686 ккал свободной энергии. Сам процесс аэробного окисления глюкозыможно разделить на 3 этапа:
1. Расщепление глюкозы до пирувата.
2. Окислительное декарбоксилирование пирувата до ацетил-КоА.
3. Окисление ацетила в цикле Кребса ( ЦТК ), сопряженное с работой цепи дыхательных ферментов.
Эти этапы можно представить также в виде общей схемы:
Глюкоза ----> 2 пируват ----->2 ацетил-КоА -----> 4СО2+ 10 Н2О | | + 2 СО | | | <--- 1 ---> | <---- 2 ----> | <---- 3 ----> |
Расщепление глюкозы до пирувата
По современным представлениям первыйэтапокисления глюкозы протекает в цитозоле икатализируетсянадмолекулярным белковым комплексом -гликолитическим метаболоном, включающим в себя до десятка отдельных ферментов.
Первый этап окисления глюкозы может быть в свою очередь разделен на 2 стадии. В реакциях первой стадии происходит фосфорилирование глюкозы, изомеризация остатка глюкозы в остаток фруктозы, дополнительное фосфорилирование уже фруктозного остатка и, наконец. расщепление гексозного остатка на два остатка фосфотриоз:
СН2ОН СН2ОРО3Н2
1. | |
С--- О С--- О
Н /Н \ОН Н /Н \ОН
С С + АТФ ------> С С + АДФ
НО\ОН Н/Н НО\ОН Н/Н
С--- С С--- С
Н ОН Н ОН
Глюкоза Глюкозо-6-фосфат
Эта реакция катализируется ферментом гексокиназой. В качестве фосорилирующего агента в клетке используется АТФ. Реакция сопровождается потерей свободной энергии порядка 5,0 ккал/моль и в условиях клетки является необратимой.
СН2ОРО3Н2
2. |
С--- О РО3Н2-О-СН2О
Н /Н \ОН | / \ Н
С С -----------> С С
НО\ОН Н/Н Н\Н НО/|
С--- С С---С СН2ОН
Н ОН ОН Н
Глюкозо-6-фосфат Фруктозо-6-фосфат
Вторая реакция, катализируемая фосфогексоизомеразой, легко обратима.
РО3Н2-О-СН2О РО3Н2-О-СН2О
| / \ Н | / \ Н
3. С С + АТФ ------> С С + АДФ
Н\Н НО/| Н\Н НО/|
С---С СН2ОН С---С СН2О-РО3Н2
ОН Н ОН Н
Фруктозо-6-фосфат Фруктозо-1,6-бисфосфат
Третья реакция катазируется ферментов фосфофруктокиназой. В этой реакции также теряется 3,4 ккал/моль энергии и она, как и гексокиназная реакция, в условиях клетки необратима.
РО3Н2-О-СН2О СН2ОН НС=О
| / \ Н | |
С С --------> С=О + НСОН
4.Н\Н НО/| | |
С---С СН2О-РО3Н2Н2СО-РО3Н2Н2СО-РО3Н2
ОН Н Фосфодигидр- 3-Фосфогли-
Фруктозо-1,6-бисфосфат оксиацетон цериновый
альдегид
Эта реакция катализируется ферментом альдолазой, реакция обратима. В результате реакции фруктозо-1,6-бисфосфат расщепляется на два триозофосфата.
СН2ОН НС=О
| --------->|
5.С=О <---------НСОН
| |
Н2СО-РО3Н2Н2СО-РО3Н2
Фосфодигидр- 3-Фосфогли-
оксиацетон цериновый
альдегид Фосфодигидроксиацетон ( ФДА ) в условиях клетки легко изомери-
зуется в 3-фосфоглицериновый альдегид ( ФГА ) при участии фермента триозофосфатизомеразыв ходе пятой реакции. Поэтому мы можем считать, что на первой стадии этого этапа затрачивается 2 АТФ, а из молекулы глюкозы образуется две молекулы 3-фосфоглицеринового альдегида.
На второй стадии первого этапа окисления глюкозы ФГА превращается в пируват. Поскольку при распаде молекулы глюкозы образуется 2 молекулы ФГА, при дальнейшем описании процесса мы дожны учесть это обстоятельство.
Следующая реакция рассматриваемого процесса является окислительной реакцией:
НС=О О=С~О-РО3Н2
6. | |
2 НСОН + 2НАД+ + 2Н3РО4 ----> 2 НСОН + 2НАДН+Н+
| |
Н2СО-РО3Н2 Н2СО-РО3Н2
ФГА 1,3-дифосфоглице-
риновая кислота
В ходе этой реакции, катализируемой дегидрогеназой 3-фосфоглице-риновогоальдегида, происходит окисление ФГА в 1,3-дифосфоглицериновую кислоту. Окисление идет путем дегидрирования, а отщепленные от субстрата атомы водорода переносятся на НАД+с образованием восстановленной формы кофермента. Энергия окисления накапли-
вается в клетке , во-первых, в виде энергии восстановленного НАДН+Н+,а, во-вторых, в виде макроэргической связи продукта окисления с участвующей в реакции фосфорной кислотой, т.е. в макроэргической связи 1,3-дифосфоглицериновой кислоты.
В седьмой реакции остаток фосфорной кислоты из 1,3-дифосфоглицерата вместе с запасом энергии, накопленной в макроэргической связи, переносится на АДФ с образованием АТФ:
О=С~О-РО3Н2 СООН
||
7. 2 НСОН + 2 АДФ ------> 2 НСОН + 2 АТФ
||
Н2СО-РО3Н2 Н2СО-РО3Н2
1,3-дифосфоглицериновая 3-фосфоглицериновая
кислота кислота
Эта обратимая реакция катализируется ферментом фосфоглицераткина-зой.
Далее идет обратимая изомеризация 3-фосфоглицериновой кислоты в 2-фосфоглицериновую кислоту при участии фермента фосфоглицератмутазы:
СООН СООН
||
8. 2 НСОН ------> 2 НСО-РО3Н2
||
Н2СО-РО3Н2 Н2СОН
В следующей , девятой по счету,реакции идет отщепление воды от 2-фосфоглицериновой кислоты:
СООН СООН
||
9. 2 НСО-РО3Н2 ------> 2 С~О-РО3Н2 + 2 Н2О
| |
Н2СОН СН2
3-фосфоглицериновая Фосфоенолпировино-
кислота градная кислота ( ФЭП )
В ходе отщепления воды идет перераспределение электронной плотности в молекуле с образованием макроэргической связи между вторым
атомом углерода енольной формы пировиноградной кислоты и остатком фосфорной кислоты. Реакция обратима, она катализируется ферментом
енолазой.
Накопленная в макроэргической связи ФЭП энергия вместе с остатком фосфорной кислоты в ходе следующей реакции переносится на АДФ с образованием АТФ. Реакция катализируется пируваткиназой.
СООН СООН
!О. ||
2 С~ О-РО3Н2+ 2 АДФ ------> 2 С=О + 2 АТФ
||
Ch3 СН3
Реакция сопровождается потерей 7,5 ккал/моль энергии и в условиях клетки практически необратима.
Суммарное уравнение первого этапа аэробного окисления глюкозы:
| Глюкоза + 2 АДФ | 2 Н3РО4+ 2 НАД+----> > 2 пируват + 2 АТФ + 2 НАДН+Н++ 2 Н2О |
| В ходе этого этапа высвобождается 140 ккал/моль энергии, ос- новная ее часть ( около 120 ккал/моль ) накапливается в клетке в виде энергии 2 АТФ и энергии 2 восстановленных НАД+ | |
из которого следует, что на первом этапе молекула глюкозы расщепляется до двух молекул пировиноградной кислоты, при этом клетка на каждую молекулу расщепленной глюкозы получает 2 молекулы АТФ и две молекулы восстановленного НАДH+H+.
Регуляция работы первого этапа аэробного расщепления глюкозы осуществляется с помощью термодинамических механизмов и с помощью механизмов аллостерической модуляции регуляторных ферментов, принимающих участие в работе этого метаболического пути.
С помощью термодинамических механизмов осуществляется контроль направления потока метаболитов по данному метаболическому пути. В описанную систему реакций включены три реакции, в ходе которых теряется большое количество энергии: гексокиназная ( G0=
- 5,0 ккал/моль ), фосфофруктокиназная ( G0= -3,4 ккал/моль ) и пируваткиназная ( G0= - 7,5 ккал/моль ). Эти реакции в клетке практически не обратимы, в особенности пируваткиназная реакция, и за счет их необратимости процесс становится необратимым в целом.
Интенсивность потока метаболитов по рассматриваемому метаболическому пути контролируется в клетке за счет изменения активности включенных в систему аллостерических ферментов: гексокиназы, фосфофруктокиназы и пируваткиназы. Таким образом, пункты термодинамического контроля метаболического пути одновременно являются и участками, на которых осуществляется регуляция интенсивности потока метаболитов.
Главным регуляторным звеном системы является фосфофруктокиназа. Активность этого фермента подавляется высокими концентрациями АТФ в клетке, степень аллостерического ингибирования фермента АТФ усиливается при высоких концентрациях цитрата в клетке. АМФ является аллостерическим активатором фосфофруктокиназы.
Гексокиназа ингибируется по аллостерическому механизму высокими концентрациями Гл-6-ф. В этом случае мы имеем делом с работой сопряженного регуляторного механизма. В клетке после угнетения активности фосфофруктокиназы высокими концентрациями АТФ накапливается Фр-6-ф, а значит накапливается и Гл-6-ф, поскольку реакция, катализируемая фосфогексоизомеразой, легко обратима. В таком случае повышение концентрации АТФ в клетке ингибирует активность не только фосфофруктокиназы, но и гексокиназы.
Очень сложно выглядит регуляция активности третьей киназы - пируваткиназы. Активность фермента стимулируется Гл-6-ф, Фр-1,6-бф
и ФГА по аллостерическому механизму - так называя активация предшественником. В свою очередь, высокие внутриклеточные концентрации АТФ,НАДН,цитрата, сукцинил-КоА и жирных кислот угнетают активность фермента по аллостерическому механизму.
В целом, расщепление глюкозы до пирувата тормозится на уровне 3 указанных киназ при высокой концентрации АТФ в клетке,т.е. в условиях хорошей обеспеченности клетки энергией. При недостатке энергии в клетке активация расщепления глюкозы достигается,во первых, за счет снятия аллостерического ингибирования киназ высокими концентрациями АТФ и аллостерической активации фосфофруктокиназы АМФ и, во-вторых, за счет аллостерической активации пируваткиназы предшественниками: Гл-6-Ф, Фр-1,6-бф и ФГА.
Каков смысл ингибирования цитратом фосфофруктокиназы и цитратом и сукцинил-КоА - пируваткиназы? Дело в том, что из одной молекулы глюкозы образуется две молекулы ацетил-КоА, который за-
тем окисляется в цикле Кребса. Если в клетке накапливаются цитрат
и сукцинил-КоА, значит цикл Кребса не справляется с окислением
уже наработанного ацетил-КоА и есть смысл притормозить его допол-
нительное образование, что и достигается ингибированием фосфоф-
руктокиназы и пируваткиназы.
Наконец, угнетение окисления глюкозы на уровне пируваткиназы при повышении концентрации жирных кислот направлено на сбережение глюкозы в клетке в условиях, когда клетка обеспечена другим, более эффективным видом энергетического топлива.
studfiles.net