
4. Аэробная реакция ресинтеза атф. Аэробные реакции это
Реакции аэробные и анаэробные - Справочник химика 21
Процесс аэробного (кислородного) и анаэробного дыхания является не только источником энергии, необходимой для осуществления разнообразных реакций, для роста и движения, но и источником образования большого количества промежуточных продуктов, которые служат материалом для синтеза. Химические превращения при аэробном дыхании схематично можно выразить в виде следующего уравнения [c.209] Строение и свойства углеводов, рассмотренные выше, позволяют понять их функции в живом организме. Будучи альдо- или кето-спиртами, т.е. уже частично окисленными органическими молекулами, углеводы легко вступают в реакции дальнейшего окисления. Вследствие этого основная функция углеводов - энергетическая углеводы различного строения являются основными поставщиками энергии и на их долю приходится более 50% всей вырабатываемой в организме энергии. В зависимости от того, какое вещество является акцептором электронов в ходе окисления, процессы окислительного распада углеводов делятся на анаэробные и аэробные (акцепторы электронов - различные химические субстраты в анаэробных процессах и кислород - в аэробных процессах). [c.77]Различают следующие основные оксидоредуктазы аэробные дегидрогеназы или оксидазы, катализирующие перенос протонов (электронов) непосредственно на кислород анаэробные дегидрогеназы, ускоряющие перенос протонов (электронов) на промежуточный субстрат, но не на кислород цитохромы, катализирующие перенос только электронов. К этому классу относят также гемсодержащие ферменты каталазу и пероксидазу, катализирующие реакции с участием перекиси водорода. [c.160]
Основным катаболическим процессом деструкции глюкозы в клетках животных и человека является последовательность ряда реакций ее окисления, в результате которьгх в анаэробных условиях глюкоза превращается в лактат, а в аэробных — в конечные продукты СО2 и воду. Ниже приведена биологическая значимость окислительных превращений глюкозы [c.242]
Зависимость анаэробного процесса от температуры та же, что и у аэробного процесса, т. е. для ее характеристики можно воспользоваться уравнением (3.16). Из-за низких скоростей реакций в анаэробных условиях сильно возрастают как время гидравлического удерживания, так и время удерживания ила в реакторе. Поэтому во многих случаях экономически выгодным становится проводить процесс при высокой температуре в реакторах меньшего объема. На рис. 3.20 представлена температурная зависимость анаэробного процесса. [c.153]
Далеко не полный перечень активируемых цинком или имеющих его в своем составе ферментов свидетельствует о зависимости реакций аэробной и анаэробной фаз дыхания от наличия цинка в тканях организма. И, очевидно, главной причиной нарушений в метаболизме при недостатке цинка является расстройство функции дыхания в целом. [c.138]
Прижизненные биохимические процессы в мышце, изучавшиеся А. В. Прлладиным, В. Энгельгардтом и М. Любимовой, Д. Фердманом, В. А. Белицером и другими советскими исследователями, связаны с физиологическим актом мышечного сокращения и заключаются в реакциях гликолиза, ресинтеза мышечного гликогена, распада и ресинтеза креатинфосфата и АТФ и изменениях сократительного белкового вещества мышцы. При этом молочная кислота, образующаяся при утомлений мышцы, в результате реакций гликолиза при отдыхе мышцы в аэробных условиях частью (около одной пятой) подвергается полному окислительному распаду, а в большей своей части превращается снова в гликоген за счет энергии реакций аэробного окисления. Одновременно с реакциями гликолиза наблюдается распад АТФ и АДФ и затем креатинфосфата, что приводит к накоплению неорганических фосфатов. При отдыхе мышцы происходит ресинтез этих соединений, требующий энергии. Таким образом, наблюдается тесная связь между реакциями анаэробного и аэробного обмена в мышце, выражающаяся в том, что в аэробных условиях в мышце анаэробный распад углеводов замедлен. [c.234]
Вообще те виды брожения, которые протекают под влиянием аэробных ферментов, сводятся к реакциям окисления анаэробные же ферменты, как, например, фермент масляной кислоты, вызывают по преимуществу реакции восстановления. [c.469]
Биологическое окисление имеет значение не только потому, что дает основное количество энергии для жизнедеятельности организма, но и потому, что освобождаемая в процессе окисления энергия одновременно локализуется в организме в форме богатых энергией соединений, при помощи которых совершаются в организме все виды полезной работы. Образование таких соединений мы уже рассмотрели при обсуждении механизма анаэробного субстратного фосфорилирования. Однако такая форма образования богатых энергией соединений в общем балансе биологического окисления не превышает 10%, и в основном их обра.зование связано с цепью реакций аэробного окисления. [c.271]
TAS в выражении ДС = АН - TAS был больше по абсолютной величине, чем член АН. AS = (ДС - ДН)/333. 18.48. ДС° равно нулю при ДЯ° - TAS = О, откуда следует, что это должно иметь место при температуре Т = = AH°/AS°. Для реакции 2(г.) -> 20 (г.) АН° = 495 кДж, AS° = 117 Дж/К. Следовательно, Г = 4230 К. Разумеется, полученное значение является лишь приближенной оценкой, в которой использованы значения АН и AS при 298 К последние вряд ли сохраняют такие значения при гораздо более высоких температурах. 18.50. Для аэробной реакции ДО = = -2875 к Дж, для анаэробной реакции ДО = = - 226 кДж. Константа равновесия первой реакции больше, она равна 3 Первая реакция позволяет получить больше энергии для выполнения работы, которая может быть израсходована на осуществление других, несамопроизвольных реакций, если они связаны с окислением глюкозы. [c.475]
Другие исследователи [39—45 ] также пытались определять скорость распада аскорбиновой кислоты в аэробных и анаэробных условиях при различных условиях проведения реакций. Однако при опытах, проведенных в анаэробных условиях, они также не учитывали следов кислорода, которые остаются в растворе и в среде инертного газа [36]. В связи с этим экспериментаторы отмечали так называемый неокислительный распад аскорбиновой кислоты, хотя в значительно меньшей степени, чем в присутствии кислорода воздуха [45]. По мнению автора, это вызвано вовлечением в цепную реакцию малых количеств кислорода. [c.242]
При доочистке применяются строго аэробные биологические пруды, необходимым условием нормальной работы которых является соблюдение оптимальных для водных организмов реакции среды, температуры, наличие растворенного кислорода (не менее 1 мг/л), постоянное перемешивание воды, препятствующее образованию застойных анаэробных зон. [c.143]
Совершенно очевидно, что азотистые соединения имеют биогенное происхождение. Весьма вероятно, что порфириновые группировки создавались еще живыми организмами и перешли в нефть в качестве унаследованного продукта. С другой стороны, источником азотистых соединений могли быть белковые йещества, потому что белки содержат до 15—19% азота. Так как белки характерны главным образом для животных организмов, именно эти последние рассматривались как исходный материал нефти. В результате распада белков образуются различные аминокислоты с одной или двумя карбоксильными группами, если распад белков происходил в анаэробных условиях. В случае аэробного разложения белков азот выделяется в виде аммиака. Анаэробное разложение белков дает кроме аминокислот некоторые циклические соединения, содержащие пироллоповые или пирролидоновые циклы. Если исходный материал нефти содержал полисахариды, возможна реакция их альдегидной группы с аминогруппой аминокислот, При этом образуются темные продукты конденсации. Этой реакции приписывается большая роль при образовании углей из смешанного целлюлозно-лигнинового материала. Продукты конденсации аминокислот с целлю лозным материалом, так называемые меланоидины, возможно, могли бы дать циклические азотистые соединения, по своему строению достаточно далекие от исходных форм. Однако все эти предположения требуют еще прямых доказательств. [c.166]
В почвах в свободной форме аммиак ННз практически не встречается. Обычно он продуцируется в небольших количествах при разложении органических удобрений. Преобладающая часть соединений азота в почвах относится к органическим в гумусовых горизонтах почв содержится до 93—99 % азота в составе органических соединений. Минеральные соединения азота помимо аммонийных представлены нитратами и нитритами. Содержание нитритов в почве невелико и составляет десятые доли миллиграмма на 1 кг почвы. Несмотря на низкое содержание, нитратам принадлежит существенная роль в реакциях превращения соединений азота в почве. Они образуются как промежуточные соединения и при нитрификации в аэробных условиях, и при денитрификации — в анаэробных. В почвенных условиях нитриты обычно малоустойчивы и при их разложении происходят, вероятно, реакции двух основных типов [c.67]
О2 (последний не участвует в осуществляемых ими метаболических реакциях), но способные расти в его присутствии, являются по типу осуществляемого ими метаболизма облигатными анаэробами, устойчивыми к О2 внешней среды. Примером таких организмов служат молочнокислые бактерии. Многие прокариоты, относящиеся к этой же группе, приспособились в зависимости от наличия или отсутствия О2 в среде переключаться с одного метаболического пути на другой, например с дыхания на брожение, и наоборот. Такие организмы получили название факультативных анаэробов, или факультативных аэробов. Представителями этой физиологической группы прокариот являются энтеробактерии. В аэробных условиях они получают энергию в процессе дыхания. В анаэробных условиях источником энергии для них служат процессы брожения или анаэробного дыхания. [c.129]
Окислительное брожение. Это такой биохимический процесс, для течения которого необходим кислород воздуха и при котором имеет место неполное использование потенциальной энергии перерабатываемого вещества (продукты реакции — органические кислоты способны к дальнейшим окислительным превращениям с выделением энергии). По первому признаку процесс окислительного брожения сходен с процессом обычного аэробного дыхания, но конечными продуктами настоящего аэробного дыхания является вода и углекислота, которые не могут дальше окисляться с выделением энергии по второму признаку он подобен настоящему брожению, но последнее является анаэробным процессом, а окислительное брожение — аэробным. Примером [c.561]
Переход от анаэробного пути распада углеводов к аэробному - пиру-ватдегидрогеназная реакция [c.82]
В клетках эубактерий компонентами липидов являются в основном насыщенные жирные кислоты или содержащие одну двойную связь (мононенасыщенные). Полиненасыщенные жирные кислоты, содержащие две и более двойных связей, найдены до сих пор только у цианобактерий. Образование двойных связей в молекуле кислоты может происходить двумя путями. Один из них, обнаруженный у аэробных эубактерий, требует участия молекулярного кислорода. У облигатно анаэробных и некоторых аэробных эубактерий двойные связи вводятся в молекулу кислоты на ранней стадии ее синтеза в результате реакции дегидратации. [c.88]
Одной из существенных черт сходства фермента и модели является предварительное образование оксигенированного комплекса. При отсутствии кислорода взаимодействие радикала с катализатором может иметь специфические особенности, что должно сказаться на направлении реакции. Действительно, если окисление 2,6-ди-трт-бутил-4-метилфенола в аэробных условиях протекает с образованием дифенола (/) и стильбен-хинона (//), то в анаэробных условиях образуется дифенохинон следующего строения [c.212]
Судьба дигидрокодегидразы изменяется в зависимости от того, происходит ли реакция в анаэробной или аэробной среде. В нервом случае KoIHj отдает водород акцептору (например, ацетальдегиду в процессе спиртового брожения), регенерируя Kol (и образуя этанол). [c.801]
Довольно противоречивы сведения о реакции отдельных экологотрофических группировок микроорганизмов на сельскохозяйственное захрязнение. Минеральные удобрения в умеренных дозах активизируют жизнедеятельность аэробных и анаэробных азотфиксаторов, денитри-фикаторов, аммонификаторов, целлюлозоразрушающих бактерий, актиномицетов, фибов, микроорганизмов автохтонной фуппировки. В то же время в длительно удобряемых дерново-подзолистых почвах внесение азотных удобрений отдельно и совместно с калийными привело к уменьшению числа азотфиксирующих, аммонифицирующих и целлюлозоразлагающих микроорганизмов. Напротив, внесение фосфорных и калийных удобрений, а также известкование влияет на повышение численности этих микроорганизмов. [c.165]
При уменьшении интервалов отдыха между упражнениями до 1 мин еще наблюдается отставленный максимум потребления О , что свидетельствует об активизации процессов восполнения алактатных анаэробных резервов с каждым очередным повторением максимального усилия. Однако он исчезает, когда продолжительность интервалов отдыха сокращается до 30 с. Вместо этого появляется пилообразная кривая с наивысшими значениями скорости потребления в конце каждого повторения максимального усилия и небольшим снижением в паузах отдыха. После первых 5—6 повторений упражнения и дальше скорость потребления не изменяется, устанавливаясь на определенном уровне, соответствующем тяжести выполняемой интервальной работы, что в данных условиях зависит от величины избранных интервалов отдыха. Если интервалы отдыха сокращаются до 10 с, уровень пикового потребления при выполнении упражнений сравнивается с величиной МПК. Сокращение интервалов отдыха в этих условиях сопровождается усилением при первых 5—6 повторениях избыточного выделения Og, быстрым накоплением молочной кислоты и снижением pH крови (рис. 186). Значительное закисление внутренних сред организма в результате накопления молочной кислоты в крови (более 10 ммоль л ) ведет к снижению скорости креатинфосфокиназной реакции и максимальной мощности. Дальнейшее увеличение числа повторений приводит к изменению тренировочного эффекта интервальной работы он приобретает смешанный аэробно-анаэробный характер. Поэтому если в интервальном методе применяются кратковременные максимальные усилия, чередуемые с короткими интервалами отдыха (менее 30 с), то для создания алактатного анаэробного эффекта тренировки такую работу следует выполнять сериями по 5—6 повторений в каждой с интервалами отдыха между сериями не менее 3 мин. [c.398]
Реакции аэробного распада углеводов катализируются теми же ферментами, которые принимали участие в анаэробном распаде фосфоферазы, дегидрогеназы, карбоксилазы и др. [c.91]
Таким образом, прп анаэробном потреблении глюкозы пз топлива, способного дать 2808 кДж/моль энергии, выделяется только 2166 кДж/моль. Позднее мы увидим, что энтальпия реакции не яв-лпстсп единственным критерием, который нсобхотимо рассматривать ири оценке эффективности использования ресурсов в процессе дыхания. Однако приведенный пример ясно показывает, что аэробное дыхание — горазю более усложненный процесс, чем более примитивная анаэробная ферментация. [c.127]
Гликолиз— это последовательность десяти ферментативных реакций, в процессе которьгх в аэробных условиях глюкоза расшепляется до двух молекул пирувата (аэробный гликолиз), а в анаэробных — до двух молекул лактата (анаэробный гликолиз). Ниже приведены стехиометрические уравнения процессов анаэробного (а) и аэробного (б) гликолиза [c.243]
При высоких значениях БПК, которые соответ-сгвуют полному исчерпанию кислорода в воде, существующие в ней аэробные бактерии (жизнедеятельность которых невозможна без кислорода) уступают место анаэробным бактериям (не требующим кислорода). В результате продукты окислительных реакций — СО2, Н2504, Н3РО4 и НКОз — сменяются продуктами восстановительных реакций — СН4, П28, РН3 и ННз. Поэтому не следует удивляться, что вода с высоким показателем БПК, например в гаванях и перегруженных сточными водами реках, имеет зловонный запах. [c.508]
Параллельно с не слишком плодотворными попытками построить обобщенную термодинамическую теорию, применимую к живым системам, проводились чисто эмпирические наблюдения над процессами роста живых систем и потребления ими энергии, выявившие ряд интересных фактов. Довольно хорошо изучены многие анаэробные процессы брожения, в ходе которых энергия химических реакций используется клетками для синтеза АТР (гл. 9). Как правило, стехиометрия этих реакций известна, и поэтому можно с хорошей точностью оценить количество АТР, синтезированного при сбраживании данного количества субстрата. Нетрудно измерить и количество образовавшейся в ходе брожения биомассы например, можно собрать культуру клеток быстро растущих бактерий, промыть, высушить и взвесить ее. Оказалось, что независимо от того, какой именно субстрат сбраживается (за редким исключением), величина Удтр — бес высушенных клеток в граммах на моль синтезированного АТР — остается почти постоянной [22, 31] и приблизительно равной 10,5. Другой факт состоит в том, что для бактерий, рост и деление которых (в аэробных условиях) сопровождается выделением только СОг и воды, 40 5% потребляемого углерода и водорода окисляется до СОа и воды, а 60 5% ассимилируется клетками. Отметим, что такой процент ассимилированного материала значительно выше, чем для анаэробного брожения, при котором подавляющая часть материала сбраживается, а не ассимилируется. Как мы увидим позднее, это различие обусловлено тем, что окисление дает значительно больший выход АТР, нежели брожение. [c.234]
При анаэробном брожении органические вещества разрущаются анаэробными бактериями. Для них, как и для аэробных бактерий, источником энергии являются окислительные процессы. Различие заключается в том, что последние получают энергию из окислительно-во с-становительных реакций, в которых акцептором водорода служит свободный кислород. Энергодающие окисли-тельно-восстановительные реакции анаэробных бактерий протекают с освобождением энергии за счет энзиматического расщепления сложных органических веществ. Подобные процессы называются бродильными. Брожение ведет к глубокому распаду веществ, но никогда не заканчивается их полным окислением. Процессы эти экзотермические, т. е. сопровождаются выделением тепла. [c.241]
Как отмечалось, одна молекула НАДН (3 молекулы АТФ) образуется при окислительном декарбоксилировании пирувата в ацетил-КоА. При расщеплении одной молекулы глюкозы образуется 2 молекулы пирувата, а при окислении их до 2 молекул ацетил-КоА и последующих 2 оборотов цикла трикарбоновых кислот синтезируется 30 молекул АТФ (следовательно, окисление молекулы пирувата до СО, и Н,0 дает 15 молекул АТФ). К этому количеству надо добавить 2 молекулы АТФ, образующиеся при аэробном гликолизе, и 6 молекул АТФ, синтезирующихся за счет окисления 2 молекул внемитохондриального НАДН, которые образуются при окислении 2 молекул глицеральдегид-З-фосфата в дегидрогеназной реакции гликолиза. Следовательно, при расщеплении в тканях одной молекулы глюкозы по уравнению gH ,Og + 60,—>6СО, + 6Н,0 синтезируется 38 молекул АТФ. Несомненно, что в энергетическом отнощении полное расщепление глюкозы является более эффективным процессом, чем анаэробный гликолиз. [c.349]
Для реализации биосинтеза и метаболизма необходима энергия, запасаемая в клетках в химической форме, главным образом в экзергонических третьей и второй фосфатной связи АТФ. Соответственно метаболические биоэнергетические процессы имеют своим результатом зарядку аккумулятора — синтез АТФ из АДФ и неорганического фосфата. Это происходит в процессах дыхания и фотосинтеза. Современные организмы несут память об эволюции, начавшейся около 3,5 10 лет назад. Имеются веские основания считать, что жизнь на Земле возникла в отсутствие свободного кислорода (см. 17.2). Метаболические процессы, протекающие при участии кислорода (прежде всего окислительное фосфорилирование при дыхании), относительно немногочисленны и эволюционно являются более поздними, чем анаэробные процессы. В отсутствие кислорода невозможно полное сгорание (окисление) органических молекул пищевых веществ. Тем не менее, как это показывают свойства ныне существующих анаэробных клеток, и в них необходимая для жизни энергия получается в ходе окислительно-восстановительных процессов. В аэробных системах конечным акцептором (т. е. окислителем) водорода служит Ог, в анаэробных — другие вещества. Окисление без Oj реализуется в двух путях брожения — в гликолизе и в спиртовом брожении. Гликолиз состоит в многостадийном расщеплении гексоз (например, глюкозы) вплоть до двух молекул пирувата (пировиноградной кислоты), содержащих по три атома углерода. На этом, пути две молекулы НАД восстанавливаются до НАД.Н и две молекулы АДФ фосфоршгируются— получаются две молекулы АТФ. Вследствие обратной реакции [c.52]
Впервые роль каротиноидов в предотвращении летального эффекта, вызываемого фотоокислением, была показана при изучении бескаротиноидного мутанта пурпурной бактерии КИоёорзеи- отопаз зркего1(1ез. Исходная культура хорошо росла фототрофно в анаэробных условиях, но могла также расти на свету и в темноте в аэробных условиях. Полученный из нее мутант, лишенный каротиноидов, обладал низкой скоростью роста на свету в анаэробных условиях и в темноте в аэробных условиях, но быстро погибал при перенесении на свет + воздух. Фотоокислительные повреждения могут развиваться и у нефотосинтезирующих прокариот, так как в их клетках также имеются окрашенные молекулы, поглощающие видимый свет, которые могут функционировать как фотосенсибилизаторы. Действие каротиноидов не ограничивается только их участием в защите от фотодинамического эффекта. Они гасят синглетное состояние кислорода независимо от того, в каких реакциях он возникает на свету или в темноте. [c.339]
Хорошо известен связанный с мембраной фермент сукцинатдегидрогеназа, катализирующий в ЦТК окисление сукцината до фумарата. Водород, акцептируемый в этой реакции флавинадениндинуклеотидом (ФАД), непосредственно поступает в дыхательную цепь (см. рис. 92). Поскольку фумаратредуктаза и сукцинатдегидрогеназа катализируют одну и ту же реакцию, но в разных направлениях, первоначально считали, что это один фермент. Сейчас показано, что реакции осуществляются разными ферментными белками. Информация о них содержится в разных генах. Синтез сукцинатдегидрогеназы индуцируется в аэробных, а фумаратредуктазы — в анаэробных условиях. [c.354]
Разделение на анаэробный и аэробный гликолиз носит условный характер, так как реакции гликолиза в присутствии кислорода и его отсутствии одни и те же. Различия касаются лишь их скорости и конечных продуктов. При недостатке кислорода реокисление НАДН, образовавшегося в ходе гликолиза, осушествляется путем сопряжения с восстановлением пирувата в лактат, а в аэробных условиях НАДН окисляется в ходе кислородзависимого процесса окислительного фосфорилирования (гл. 15), результатом которого является образование большого количества АТФ. [c.243]
Массообмен в биологаческих системах связан не только с доставкой кислорода к клеткам аэробных видов и теплообменом, но и с выделением диоксида углерода как конечного продукта различных катализируемых реакций, транспортом других веществ через клеточные мембраны (в том числе — анаэробных организмов) [c.266]
Газообразные отходыв процессах биологической технологии немногочисленны в ассортименте. Это определяется биохимической сущностью реакций, катализируемых ферментами. Как правило, энергетическим субстратом для биообъектов являются углеводы. В аэробных и анаэробных условиях из них образуется диоксид углерода. Так, при брожении (гликолитический процесс) из глюкозы образуется 2 моля СО2 [c.365]
Окислительно-восстановительный потенциал. Жизнедеятельность бактерий зависит от потенциала. Процессы превращения остатков органических соединений при свободном доступе кислорода (аэробные условия) и воды аналогичны медленному горению и называются тлением. Гумификация (перегни-вание) характеризуется недостаточным доступом воздуха (анаэробные условия) и влаги. Этот процесс приводит к накоплению зауглероженпого остатка (гумуса), часть которого может растворяться в воде. Превращение органических веществ в условиях избытка влаги и отсутствия кислорода широко распространено в природе и называется оторфением оно приводит к появлению твердых гумусовых продуктов. Образование сап-ропелей из водорослей и планктона протекает в отсутствие кислорода под слоем воды (восстановительные реакции) и известно как процессы гниения, или гнилостного брожения. [c.30]
Нами было показано, что комплекс, образующийся нри оксигениро-вании солей одновалентной меди в растворе пиридина, во многих отношениях аналогичен ферменту тирозиназе [25]. И в ферменте, и в модельном катализаторе в анаэробных условиях медь находится в одновалентном состоянии в аэробных условиях происходит оксигенирование, причем на два атома меди поглощается один моль О 2- Стадия образования оксигенированного комплекса предшествует стадии окисления. В аэробных условиях часть меди сохраняется в одновалентном состоянии. Из двух атомов молекулы кислорода фермент и его модель переносят к субстрату только один. Субстрат восстанавливает часть меди до Си . Перекись водорода не участвует в процессе. Образуется промежуточный радикал, стабилизированный на ферменте или модели. Для фермента были предположены координационные связи Си—N и показано отсутствие геми-новоп структуры, для модели методом ЭПР найдена связь u N фта-лоцианин меди не является катализатором реакций, проводимых моделью. [c.210]
Колиформная группа бактерий определяется как совокупность аэробных и факультативно-анаэробных, неспорообразующих, грамотри-цательных палочек, вызывающих брожение лактозы с образованием газа в течение 48-часовой инкубации при 35°С. Первоначальный анализ на колиформы — предположительный, он основан на выделении газа из лактозы. Десятимиллилитровые порции водной пробы переносят с помощью стерильных пипеток в подготовленные бродильные трубки. Трубки содержат лактозу или лауриновую триптозу, бульон и перевернутые ампулы (сборники газа). Инокулированные трубки помещают в термостат с температурой 35 0,5°С. Рост бактерий с выделением газа, обнаруживаемого по появлению пузырьков в перевернутой ампуле, считается положительным анализом, показывающим, что колиформные бактерии могут присутствовать. Отрицательная реакция (отсутствие роста или рост без газа) исключает присутствие бактерий колиформной группы. [c.66]
chem21.info
Аэробные и анаэробные нагрузки | ТХЭКВОНДО В ЧЕЛЯБИНСКЕ
В реальном боевом столкновении победу над противником может одержать человек, обладающий весьма скромными физическими данными. Например, благодаря умелому применению жестоких травмирующих приемов или оружия.
Но в спортивных поединках одним из важнейших условий достижения победы, помимо технического или тактического превосходства, является соответствующая физическая кондиция («форма»).
Первый и самый важный ее компонент, это скоростно-силовая выносливость. Речь идет о способности спортсмена тренироваться с такой интенсивностью, когда частота сердечных сокращений (чсс) находится в интервале 130—190 в минуту, так как только при этом достигается наибольший тренировочный эффект. Тренировочные нагрузки с показателем чсс менее 130, малоэффективны для тренировки аэробных возможностей организма.
Аэробные реакции — это основа биологической энергетики. Источник биологической энергетики при мышечной работе — АТФ (аденозинтрифосфорная кислота). Заменить АТФ другими источниками энергии невозможно. Чем больше спортсмен потребляет кислорода в единицу времени, тем большее количества АТФ образуется в мышцах. По этой причине максимальное потребление кислорода (МПК) является самым информативным показателем уровня аэробных возможностей спортсмена.
В анаэробном виде двигательной деятельности энергия вырабатывается за счет быстрого химического распада «топливных» веществ в мышцах без участия кислорода. Этот способ срабатывает мгновенно, но быстро истощает запасы готового «топлива» (0,5 — 1,5 мин), после чего запускается механизм аэробной выработки энергии
Характерные примеры анаэробной двигательной деятельности — силовая подготовка и спринтерский бег. Различия между двумя типами двигательной деятельности происходят от разной продолжительности и интенсивности мышечных сокращений. От этого зависит способ, которым энергия производится внутри мышц.
Первоначально при повышенной нагрузке мышечный гликоген перерабатывается в глюкозу в процессе гликолиза, образуя пируват, который после этого реагирует с кислородом (цикл Кребса), чтобы произвести углекислый газ и воду, выделяя энергию. При нехватке кислорода (например при выполнении взрывных движений, которые являются анаэробными упражнениями), углеводы потребляются быстрее, так как пируват метаболизируется до лактата. Когда количество углеводов истощается, метаболизм жиров повышается для создания топлива через метаболические пути аэробного гликолиза. Анаэробные упражнения часто относятся к начальной фазе двигательной деятельности, приходящуюся на начало физической нагрузки или происходящую во время любых резких рывков интенсивной нагрузки. При нагрузке такой интенсивности гликоген используется без участия кислорода и этот процесс менее эффективен.

Таблица Фокса-Хаскела показывает зависимость между аэробной (светло-оранжевый) и анаэробной (темно-оранжевый) нагрузкой и пульсом.
Аэробными являются очень многие виды физических упражнений. Например, бег на дальние дистанции в среднем темпе — характерный пример аэробной нагрузки, а спринтерский бег на короткой дистанции — анаэробной. Игра в теннис между двумя участниками, состоящая из плавных и постоянно повторяющихся движений — в основном аэробная нагрузка, в то время как гольф или командный теннис, состоящие из резких всплесков нагрузки, включают большую часть анаэробной. Существуют аэробные виды спорта по своей природе, а кроме того, разработаны специальные упражнения с максимальной аэробной составляющей — фартлек, аэробика.
Преимущества, которые дает регулярная аэробная тренировка:
- укрепляются мышцы, ответственные за дыхание;
- укрепляется сердечная мышца, увеличивается её эффективность, снижается пульс в состоянии покоя;
- укрепляются скелетные мышцы во всем организме;
- улучшается циркуляция крови, снижается кровяное давление;
- увеличивается число красных кровяных телец, доставляющих кислород в ткани;
- улучшается психическое состояние, уменьшается стресс, снижается риск депрессии;
- снижается риск диабета .
Эффект от тренировок проявляется лишь в том случае, когда человек выполняет их с достаточной интенсивностью и достаточно часто. Чаще всего рекомендуют не менее 20 минут три раза в неделю.
Аэробные упражнения не дают столь же значительной прибавки в физической силе, как анаэробные. Поэтому для профессионалов — спортсменов, военнослужащих, пожарной охраны и полиции, необходимо совмещение обоих типов тренировок. Механизм питания мышц под большой и резкой нагрузкой может быть развит только с помощью анаэробных тренировок. Тем не менее, аэробные упражнения вносят отличный вклад в развитие сердечно-сосудистой системы, необходимое для выносливости.
tkd174.ru
Переключение между аэробным и анаэробным окислением происходит автоматически
Прекрасной иллюстрацией способности метаболизма переключаться с одного направления на другой без какого-либо дополнительного вмешательства является эффект Пастера.
Эффект Пастера
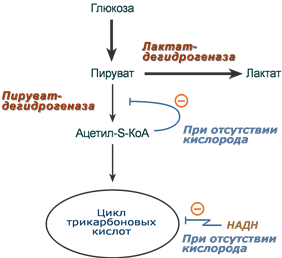
Эффект Пастера - это снижение потребления глюкозы и прекращение продукции молочной кислоты клеткой в присутствии кислорода. Биохимическая основа эффекта заключается в конкуренции за субстрат между пируватдегидрогеназой, превращающей пируват в ацетил-S-КоА, и лактатдегидрогеназой, превращающей пируват в лактат.
У пируватдегидрогеназы сродство гораздо выше и в обычных аэробных условиях она окисляет большую часть пировиноградной кислоты. Как только поступление кислорода уменьшается (недостаток кровообращения, спазм сосудов, тромбозы и т.п.) происходит следующее:
В этой ситуации пировиноградной кислоте не остается ничего иного как превращаться в молочную.
При наличии кислорода ингибирование ПВК-дегидрогеназы прекращается и она, обладая большим сродством к пирувату, выигрывает конкуренцию.
Отличной иллюстрацией к сказанному служит чувствительность миокарда и нейронов к недостатку кислорода:
Роль лактатдегидрогеназы в клетке
В норме работу клеток нервной системы и миокарда при аэробных условиях обеспечивает большое количество митохондрий и поступление из крови субстратов для окисления – глюкозы, кетоновых тел, жирных кислот (только для миокарда), лактата, превращегося в пируват.
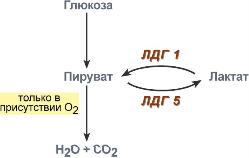 В основе высокой чувствительности этих органов к отсутствию кислорода лежит отличие изоферментов лактатдегидрогеназы друг от друга. Рассмотрим это на прмиере ЛДГ-1 и ЛДГ-5.
В основе высокой чувствительности этих органов к отсутствию кислорода лежит отличие изоферментов лактатдегидрогеназы друг от друга. Рассмотрим это на прмиере ЛДГ-1 и ЛДГ-5.
Сердечный изофермент ЛДГ-1, обладая высоким сродством к молочной кислоте, переводит ее в пировиноградную, "стремясь" поднять концентрацию пирувата с целью его включения в цикл трикарбоновых кислот (только в аэробных условиях) и получения энергии для сокращения миокардиоцита. Поэтому миокард не образует лактат, но зато может использовать его для получения энергии, захватывая из крови, но только в аэробных условиях. Аналогично лактат используется нервной системой.
При нехватке кислорода свойства изофермента ЛДГ-1 не изменятся, он по-прежнему будет сдвигать реакцию в сторону продукции пировиноградной кислоты (на схеме "справа-налево"). Однако без кислорода "сгореть" в ЦТК пируват не может и выйти из клетки тоже не может, т.к. клеточные мембраны непроницаемы для него. Являясь кислотой, он закисляет цитозоль, изменяя активность ферментов и, возможно, как-то еще проявляет токсичность. Таким образом, происходят необратимые повреждения кардиомиоцита и нейрона – развивается инфаркт миокарда. или ишемический инсульт.
В тоже время изофермент скелетной мышцы ЛДГ-5 обладает высоким сродством к пирувату, при отсутствии кислорода в клетке быстро и эффективно превращает его в молочную кислоту, обратная же реакция практически не идет. Закисление саркоплазмы снижает работоспособность миоцита. Мышца, хоть и не в состоянии работать в таких условиях, все-таки сохраняет жизнеспособность. Но в клетке молочная кислота не накапливается – она легко проходит через клеточные мембраны и быстро удаляется из мышцы. После "отключки" скелетная мышца вновь становится работоспособной через несколько минут.
Таким образом, в анаэробных условиях сильнее всего будут страдать сердечная мышца и нервная система, что, собственно говоря, и наблюдается в медицинской практике.
Вы можете спросить или оставить свое мнение.
biokhimija.ru
4. Аэробная реакция ресинтеза атф
1. Дыхательное фосфорилирование
При более длительной (больше 2-3 мин.), но менее интенсивной работе (например, умеренной мощности – бег на длинные и сверхдлинные дистанции) мышечные клетки все в большей степени обеспечиваются достаточным количеством кислорода. Поэтому ресинтез АТФ из АДФ все в большей мере будет осуществляться за счет дыхательного фосфорилирования, т.е. благодаря взаимосвязи (сопряжения) аэробного окисления или дыхания (например, глюкозы, молочной кислоты, жирных кислот) – с процессом фосфорилирования АДФ. В конечном итоге образуется 38 молекул АТФ (чистый выход – 36 молекул АТФ) (3.6).
Наибольшей скорости дыхательное фосфорилирование достигает к 5-6 минуте работы и мало изменяется при большей ее продолжительности.
Общий выход энергии при дыхательном фосфорилировании – в 10 раз превышает гликолиз и на его долю приходится около 90% от общего количества АТФ, ресинтезируемого в клетках организма. Мощность аэробных реакций ресинтеза АТФ оценивается по величине максимального потребления кислорода (МПК), доступного для выполнения мышечной работы.
При работе большой интенсивности содержание АТФ в мышечных клетках резко снижается, в результате чего происходит набухание митохондрий, внутри которых увеличивается расстояние между отдельными гребнеподобными мембранами. В таких случаях наблюдается разобщение между аэробным окислением и фосфорилированием, вследствие чего резко снижается ресинтез АТФ и увеличивается теплообразование, что способствует увеличению температуры тела. По мере улучшения снабжения мышц кислородом – нормальное соотношение между реакциями аэробного окисления и фосфорилирования восстанавливается и частичное разобщение устраняется. Такое явление наблюдается во время энергичной разминки («разогревания»), при выполнении упражнений большой интенсивности, а также при сильном утомлении.
5. Соотношение процессов аэробного и анаэробного ресинтеза атф при мышечной работе разной мощности и длительности
Независимо от характера и длительности мышечной работы, источником энергии всегда является расщепление АТФ. Однако, чтобы работа продолжалась во времени, необходимо восстановление (ресинтез) АТФ. Участие различных реакций ресинтеза АТФ в динамике мышечной деятельности можно представить графически (3.7).
Из графика следует, что в первые 2-3 сек. работы – работа осуществляется за счет расщепления АТФ. С 3 по 20 секунду – ресинтез АТФ обеспечивается алактатной КФКР. С 30 по 40 сек. – происходит максимальная интенсификация ресинтеза АТФ за счет гликолитического фосфорилирования (гликолиза). В дальнейшем происходит постепенное, все в большей степени, превалирование дыхательного фосфорилирования (например, аэробного окисления глюкозы).
Таким образом, в начале всякой работы, а при работе максимальной и субмаксимальной интенсивности, на всем ее протяжении, ресинтез АТФ осуществляется анаэробным путем: сначала за счет КФ, а затем за счет гликолиза. По мере продолжения работы – гликолиз постепенно сменяется аэробным окислением.
В зависимости от интенсивности работы наблюдается то или иное соотношение анаэробного и дыхательного фосфорилирования. Установлено, что если общая доля участия алактатного и гликолитического анаэробных процессов при выполнении упражнений составляет 60 % от энергетического запроса, то такие упражнения называются упражнениями анаэробного характера. Если аэробный процесс составляет 70 % и более – это упражнения аэробного характера. Если аэробные и анаэробные процессы имеют равное значение, то такие упражнения называются упражнениями аэробно-анаэробного (т.е. смешанного) характера.
studfiles.net
Анаэробная реакция
Анаэробное дыхание — реакции распада глюкозы без участия кислорода.[ ...]
Реакция протекает в анаэробных условиях.[ ...]
Реакция восстановления нитратов до нитритов идет в анаэробных условиях, поэтому воздух из трубок вновь выкачивают в течение 5 мин. Затем выдерживают в ванне термостата при 37 °С 30 мин. После этого реакцию останавливают, перелив уксусную кислоту из шарика в реакционную среду. Если раство р мутный, в него для осаждения белков добавляют 3 мл насыщенного раствора (ЫН4)2504.[ ...]
Анаэробные бактерии живут в воде, лишенной кислорода, или в иле, где формируется восстановительная среда, и вырабатывают метан в процессе разложения углеводородов простой структуры. В такой же среде встречаются и некоторые виды сульфобактерий, которые восстанавливают сульфаты сернистых соединений и вырабатывают сероводород. Эти микроорганизмы способствуют образованию черного сернистого железа в результате реакций железа с сероводородом, и поэтому грязь и ил окрашены в характерный черный цвет.[ ...]
Анаэробные процессы используются в практике при обеззараживании и разложении осадков сточных вод. Из-за малого теплового эффекта этой реакции анаэробный процесс является удобным для санитарной техники, так как при синтезе микробной клетки перерабатывается значительное количество органического вещества.[ ...]
Анаэробная ферментация органического материала применяется при обработке бытовых и промышленных стоков, а также отходов животноводческих и птицеферм [381, 404]. Хотя анаэробное сбраживание органического материала в метантенках используется уже давно, бактериология и биохимия этого процесса изучены недостаточно. Одной из причин этого, по-видимому, являются трудности, с которыми встречаются исследователи при культивировании анаэробных бактерий [381]. Успехи в изучении микрофлоры, участвующей в анаэробном разложении отходов, были достигнуты после получения новых сведений о микроорганизмах рубца жвачных животных. Процессы, протекающие в рубце, имеют много общего с реакциями превращения органических веществ в метантенках. Вот почему исследования, которые ведутся в этих двух направлениях, взаимосвязаны [380].[ ...]
Анаэробные организмы не нуждаются в молекулярном кислороде и получают энергию за счет окислительно-восстановительных внутримолекулярных реакций. Процессы освобождения энергии протекают под влиянием комплекса дегидрирующих ферментов и называются бродильными. Образующиеся при этом соединения используются в реакциях синтеза для построения органического вещества клетки.[ ...]
В анаэробных условиях и при кислой реакции среды количество подвижного марганца в почвах сильно возрастает. Поэтому на кислых заболоченных почвах растения иногда страдают от избытка марганца. Токсичность марганца на этих почвах устраняется внесением извести, — в этом случае марганец переходит в менее подвижное состояние.[ ...]
При анаэробном брожении органические вещества разрушаются анаэробными бактериями. Для них, как и для аэробных бактерий, источником энергии являются окислительные процессы. Различие заключается в том, что последние получают энергию из окислительно-восстановительных реакций, в которых акцептором водорода служит свободный кислород. Энергодающие окисли-тельно-восстановительные реакции анаэробных бактерий протекают с освобождением энергии за счет энзиматического расщепления сложных органических веществ. Подобные процессы называются бродильными. Брожение ведет к глубокому распаду веществ, но никогда не заканчивается их полным окислением. Процессы эти экзотермические, т. е. сопровождаются выделением тепла.[ ...]
Кислая реакция стоков и большое содержание органических веществ вызывают развитие плесени. Затрудняется доступ кислорода к фильтрующему материалу, приостанавливается минерализация, в толще биофильтра аэробные процессы сменяются анаэробными, появляется неприятный запах, очистка прекращается.[ ...]
Зависимость анаэробного процесса от температуры та же, что и у аэробного процесса, т. е. для ее характеристики можно воспользоваться уравнением (3.16). Из-за низких скоростей реакций в анаэробных условиях сильно возрастают как время гидравлического удерживания, так и время удерживания ила в реакторе. Поэтому во многих случаях экономически выгодным становится проводить процесс при высокой температуре в реакторах меньшего объема. На рис. 3.20 представлена температурная зависимость анаэробного процесса.[ ...]
Эти отдельные реакции протекают в сотые доли секунды (второстепенные реакции в этой схеме опущены). Для начала гликолиза требуется энергия, получаемая путем разложения двух частиц АТФ. За счет ее происходит разложение фруктозы на две части. Образующиеся при этом последовательно две частицы фосфоглицеринового альдегида легко окисляются (путем выделения водорода) имеющимися в спермиях ферментами в двуфосфоглицери-новую кислоту. Получающаяся при окислении энергия служит для образования четырех частиц АТФ из аденозиндифосфата, а двуфосфоглицериновая кислота при отпадении фосфатного радикала превращается сначала в фосфо-пировиноградную, затем пировиноградную и, наконец, в молочную кислоту. Для образования последней водород, выделенный ранее из фосфоглицериио-вого альдегида, переносится на пировиноградную кислоту при помощи особого кофермента НАД, превращая ее в молочную. Из освобождающихся при этих реакциях 50 ккал энергии 33 ккал откладываются в виде АТФ, а часть энергии рассеивается в виде тепла. Сперма от образующейся молочной кислоты становится более кислой. Первой обычно расходуется глюкоза, затем фруктоза. Имеющийся в сперме быка и барана сахар, как правило, не расходуется до конца, так как образующаяся молочная кислота (довольно сильная) термозит гликолиз. В анаэробных условиях процент превращения сахара в молочную кислоту достигает в сперме быка 82,6, в аэробных — 56,1.[ ...]
Существование общего предшественника ацетата и ацетоина типа «активного ацетальдегида» предположил Schweet еще в 1951 г. [29]. Убедительные экспериментальные даказательства образования одного и того же промежуточного соединения при окислительном декарбоксилировании, катализируемом ПД из сердца свиньи, и неокислительном декарбоксилировании, осуществляемом пируватдекарбоксилазой из дрожжей, было дано Holzer в 1961 г. [9]. Рассматривая возможные превращения промежуточного продукта в различных ферментных системах, требующих для своего действия ТДФ, автор пришел к выводу, что ОЭТДФ является общим промежуточным соединение при любых превращениях а-кетокислот, связанных с их декарбоксилированием (схема 4).[ ...]
Противоположная реакция длиинодневных и короткодневных видов на анаэробиоз в период дополнительного света видна нэ рис. 67 и 68: анаэробиоз задержал цветение рудбекии и ускорил образование метелок у проса на длинном дне. Следовательно, анаэробиоз нарушает нормальное прохождение фотохимических реакций во второй половине длинного дня, что, в свою очередь, задерживает цветение длиинодневных видов и ускоряет цветение короткодпевных в условиях длиннодневного фотопериодического цикла. Этп опыты показали, что фотохимические реакции дополнительного периода света длиннодпевиого цикла являются реакциями окислительными, так как их течение резко нарушается при помещении растений в анаэробные условия.[ ...]
У аэробных бактерий анаэробные условия выращивания подавляют как спорообразование, так и рост. Спорообразование аэробных бактерий протекает лишь в условиях доступа кислорода. Интенсивность споруляции подавляется в разной степени при выращивании бактерий в неблагоприятных условиях — при температуре выше оптимальной или реакции среды менее подходящей для обильного развития. Неблагоприятные условия роста, ускоряющие споруляцию бактерий, как правило, не обусловливают обильного «урожая» спор.[ ...]
Определим, какие химические реакции лежат в основе пере-гнивания ила, если возраст ила в анаэробном осадке равен 20 суткам, а акцептором является С02. Используем для этого полуреакцию для бытовых сточных вод, содержащих ил.[ ...]
Под адаптациями к гипоксии понимаются реакции, протекающие на тканевом уровне и обеспечивающие устойчивость к недостаточному снабжению тканей кислородом. Сюда относится неспецифическая реакция повышения тканевой устойчивости к гипоксии, а также усиление анаэробного гликолиза, позволяющего осуществлять энергетические процессы при недостаточном поступлении кислорода к тканям.[ ...]
Аммонификация осуществляется аэробными и анаэробными микроорганизмами, которые составляют обширные группы бактерий, актиномицетов и плесневых грибов. Из них типичными представителями бактерий являются: Bact. vulgare, Bact. putidum, Bact. subtilis, Bact. mesentericus, Bact. micoides; представителями плесневых грибов — Aspergillus, Pénicillium, Tri-choderma koningi. Аммиак образуется во всех почвах при разной реакции среды, в присутствии воздуха и без него. На скорость аммонификации оказывают действие температура почвы, влажность и другие факторы.[ ...]
Свежая сточная жидкость имеет слабо щелочную реакцию. В результате анаэробных процессов в сточной жидкости и в иле могут образоваться органические кислоты, которые нейтрализуются бикарбонатами и карбонатами воды. Однако, по мере истощения щелочного резерва воды, реакция может стать кислой и pH ниже 7,0. Очищенная в аэробных условиях сточная жидкость имеет pH около 7,3. Активная реакция (pH) сточной жидкости имеет большое влияние на биохимические процессы, связанные с жизнедеятельностью разных групп микроорганизмов, которые относятся различно к данной концентрации водородных ионов. Кроме того, pH имеет значение для процесса биохимического коагулирования органических коллоидов и осаждения тонко диспергированной взвеси, для процесса созревания и распада ила, а также для его обезвоживания. Вследствие этого величина pH воды на различных стадиях очистки сточных вод является показателем, дающим возможность своевременно реагировать на всякое отклонение от нормального хода процесса очистки.[ ...]
Для ртути и олова было обнаружено, что в морских анаэробных условиях, т.е. в отложениях отмерших водорослей, они, присоединяя водород, переходят в летучие соединения. Такого рода гидрирование может осуществляться и с другими тяжелыми металлами. Эти реакции показывают, что на отдельных участках моря, покрытых обильной растительностью, покров из водорослей ие только угрожает морским обитателям, но и повышает «активность» тяжелых металлов-, переводя их в форму летучих гидридов, в которой они покидают воду.[ ...]
Разложение бытовых отходов, как правило, происходит в анаэробных условиях, без доступа воздуха. В результате биохимических реакций выделяются зловонные газы и пары (сероводород, аммиак, индол, скатол, метилмеркаптан), способные токсически воздействовать на организм человека.[ ...]
В соответствующих пунктах настоящей работы описаны особенности анаэробной и аэробной стабилизации осадков, методы обезвоживания, обеззараживания и обезвреживания осадков. Наряду с указанными методами, как было уже сказано выше, стабилизация осадков достигается изменением активной реакции среды.[ ...]
Брожением называется такой процесс, при котором ферментативная окислительно-восстановительная реакция проходит в анаэробных условиях при участии одного соединения, а акцептором водорода является один из продуктов реакции. Чаще всего брожение наблюдается при разложении углеводов.[ ...]
Как уже указывалось, многие группы бактерии (например, факультативные анаэробы) способны и к аэробному, и к анаэробному дыханию, но конечные продукты этих двух реакций различны и количество высвобождающейся энергии при анаэробном дыхании значительно меньше. На рис. 2.7 представлены результаты интересного исследования, в котором один и тот же вид бактерий, АегоЪас1ег, выращивали в анаэробных и аэробных условиях с использованием глюкозы в качестве источника углерода. В присутствии кислорода почти вся глюкоза превращалась в бактериальную биомассу и СО2; в отсутствие же кислорода разложение было неполным, гораздо меньшая часть глюкозы превращалась в углеродсодержащие вещества клеток и во внешнюю среду выделялся ряд органических соединений. Чтобы окислить их, потребовались бы другие специализированные виды бактерий. Когда скорость поступления органического детрита в почву и донные отложения высока, бактерии, грибы, простейшие и другие организмы создают анаэробные условия, используя кислород быстрее, чем он диффундирует в субстрат. При этом разложение органического вещества не прекращается — оно продолжается, хотя часто в замедленном темпе, если в среде имеются микроорганизмы с достаточно широким диапазоном анаэробных типов метаболизма.[ ...]
В 1970 г. Stadtman и др. сообщили о йовой В -зависимой реакции изомеризации а-метиленглу-Таравой кислоты в диметилмалеиновую кислоту [38]. Эти две кислоты являются промежуточными продуктами анаэробного сбраживания никотиновой кислоты до пропионата, ацетата, С02 и Nh4 у Clostridium. Предполагаемый механизм этой реакции представлен- ниже. Водород от С-5 а-метиленглутаровой кислоты перемещается к С-4, а остаток акрилата (углероды 2, 3 и 7) — к С-5. Перемещение водорода от С-5 к С-4 зависит от ДБК-кофермента. Перемещение двойной связи, которое должно иметь место при образовании диметилмалеииовой кислоты, требует второй протон-переносящей реакции, природа которой еще неизвестна. Как видно из предложенной схемы, эта реакция изомеризации подобна мутазным превращениям, описанным выше.[ ...]
Причинами, нарушающими работу аэротенка, являются: перегрузка очистного сооружения органическими веществами, образование анаэробных зон, недостаток биогенных элементов, резкое изменение температуры или активной реакции среды, попадание в очищаемую воду токсических веществ и т. д.[ ...]
Осадок сточных вод и концентрированные производственные сточные воды с ВПК выше 5 г/л подвергаются биохимическому разложению в анаэробных условиях. Оно может происходить в сооружениях-септиках, представляющих собой отстойник, через который медленно проходит сточная жидкость. В двухъярусном отстойнике осадок отделен от проходящей сточной жидкости, его разложение осуществляется в иловой камере. На очистных сооружениях большой производительности осадок сточных вод выделяется в первичных отстойниках и вместе с избыточным активным илом подвергается сбраживанию в метантенках. Интенсивность и глубина разложения осадка прежде всего определяются его составом, который колеблется по соотношению содержания основных органических компонентов (углеводов/ белков, жироподобных соединений) и неорганических веществ. Обычно в осадке городских сточных вод содержится 70—80% органических веществ. Так, примерный состав осадка (%): белки 24, углеводы 23, жироподобные вещества до 30. Чаще всего при кислом брожении осадка получаются уксусная, масляная, пропионовая кислоты. Образующиеся газы содержат диоксид углерода, метан, водород, сероводород. Водная фаза имеет кислую реакцию среды (pHС5), не обладает буферными свойствами, имеет резкий неприятный запах.[ ...]
Дестабилизация процесса нитрификации нарушает поступление в биологический круговорот нитратов, количество которых предопределяет ответную реакцию на изменение среды обитания у комплекса денитрификаторов. Ферментные системы денитрификаторов уменьшают скорость полного восстановления, слабее вовлекая закись азота в конечный этап, осуществление которого требует значительных энергетических затрат. В результате этого содержание закиси азота в надпочвенной атмосфере эродированных экосистем достигало 79 - 83% (Косинова и др., 1993). Отчуждение части органических веществ из черноземов под воздействием эрозии отражается на пополнении азотного фонда в ходе фото- и гетеротрофной фиксации азота: аэробной и анаэробной. На первых этапах эрозии быстрыми темпами идет подавление именно анаэробной азотфиксации в силу параметров лабильной части органического вещества (Хазиев, Багаутдинов, 1987). Активность ферментов инвертазы и каталазы в сильносмытых черноземах по сравнению с несмытыми уменьшилась более чем на 50%. В серых лесных почвах по мере увеличения их смытости наиболее резко снижается инвертазная активность. Если в слабосмытых почвах отмечается постепенное затухание активности с глубиной, то в сильносмытых уже в подпахотном слое инвертазная активность очень мала или не обнаруживается. Последнее связано с выходом на дневную поверхность иллювиальных горизонтов с крайне низкой активностью фермента. По активности фосфатазы и, особенно, каталазы четко выраженной зависимости от степени смытости почв не наблюдалось (Личко, 1998).[ ...]
При доочистке применяются строго аэробные биологические пруды, необходимым условием нормальной работы которых является соблюдение оптимальных для водных организмов реакции среды, температуры, наличие растворенного кислорода (не менее 1 мг/л), постоянное перемешивание воды, препятствующее образованию застойных анаэробных зон.[ ...]
Практически неисчерпаемый резервуар атмосферного молекулярного азота недоступен для подавляющего большинства ждавых существ. Биологическая фиксация азота осуществляется очень специализированной группой анаэробных бактерий, населяющих корневые клубеньки бобовых растений. С помощью особого фермента эти почвенные микробы осуществляют реакцию, для которой при промышленной фиксации азота требуется дорогой катализатор, температура 500° и давление до 1000 атмосфер. Некоторое количество молекулярного азота окисляется до N0 при грозовых разрядах и фотохимических реакциях в атмосфере.[ ...]
Корни многих высших растений в подтопленную почву не прорастают; если же зеркало подпочвенных вод поднимается уже после того, как корни проникли в глубь почвы, то они отмирают. Явления эти могут отчасти представлять собой прямую реакцию на нехватку кислорода, а отчасти — реакцию на накопление некоторых газов (таких, например, как сероводород, метан и этилен), выделяемых в результате жизнедеятельности микроорганизмов, участвующих в процессах анаэробного разложения органических веществ. Даже если корни не отмирают при нехватке кислорода, они могут прекратить всасывание минеральных веществ и растения все равно будут страдать от минерального голодания.[ ...]
АЭРбБНЫЕ ОРГАНЙЗМЫ - организмы, способные существовать только при наличии свободного кислорода. К А. о. относятся почти псе животные и растения, а также многие микроорганизмы, которые используют для жизнедеятельности энергию, освобождающуюся при реакциях окисления, идущих с поглощением свободного кислорода.[ ...]
Круговорот серы. Данный цикл охватывает воду, почву и атмосферу. Основные резервы серы находятся в почве и в отложениях. Содержание серы в воздухе относительно невелико. На рис. 3.5 представлен этот круговорот, ключевым звеном которого являются процессы аэробного окисления сульфида (сероводорода) до сульфата (804 ) и анаэробного восстановления сульфата до сульфида. Эти реакции осуществляются соответствующими группами бактерий. Благодаря окислительно-восстановительным процессам происходит обмен серы между фондом доступного сульфата в аэробной зоне почвы и фондом сульфидов железа, расположенным глубоко в почве и в осадках (в анаэробной зоне). В результате микробного восстановления глубоководных отложений к поверхности воды движется НгЭ, что, например, типично для Черного моря. Выделяющийся из воды сульфид окисляется до сульфата атмосферным кислородом.[ ...]
Как уже было отмечено выше, мембраны клеток состоят из гликолипидов, липидная часть которых состоит из остатков высших жирных кислот. Поэтому вероятность проникновения во внутрь клеток жирных кислот и их солей весьма высока ("подобное растворяется в подобном"). Таким образом, жирные кислоты способны ингибировать основной источник жизнедеятельности клеток.[ ...]
| Круговорот серы, охватывающий воздух, воду и почву. «Кольцо» в центре схемы иллюстрирует процессы окисления (О) и восстановления (Б), благодаря которым происходит обмен серы между фондом доступного сульфата (804) и фондом сульфидов железа, находящимся глубоко в почве и в осадках. Специализированные микроорганизмы выполняют следующие реакции | 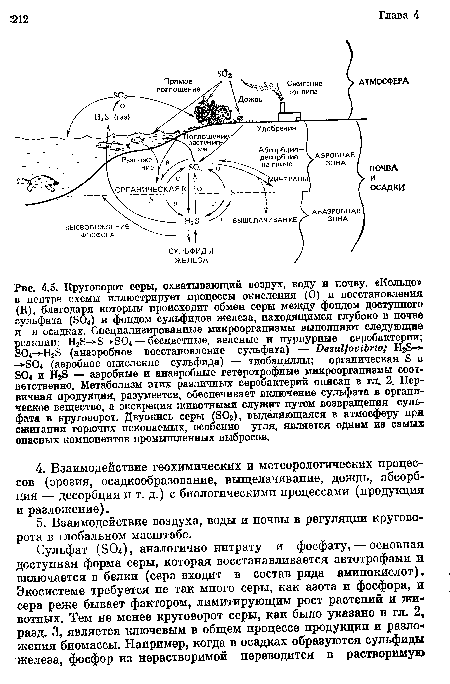 |
Проведена сравнительная оценка унифицированного метода, в котором используют железо-сульфитную среду Вильсона — Блера без антибиотиков и температуру инкубации 37°С, и нашей модификации с использованием элективной модифицированной среды СПИ и температуры инкубации 44—45°С. После подсчета черных колоний, вырастающих в том и другом случае, каждую из них идентифицировали по реакции на лакмусовом молоке, по спорообразованию и морфологии клеток. Сравнительная оценка методов выполнена при исследовании воды водоема в процессе самоочищения и на этапах очистки питьевой воды по сезонам года. В зимний период существенной разницы между индексами клостридий, определенных изучаемыми методами, не получено. В летний период черные колонии, вырастающие при 37°С, на 90— 99% состоят из гнилостных анаэробных палочек и кокков, редуцирующих сульфит, не являющихся прямыми показателями фекального загрязнения. Совместный учет этих сапрофитных бактерий с клостридиями значительно искажает результаты, вследствие чего эта группа теряет санитарно-показательное значение.[ ...]
Из коллоидных и ионных растворов, содержащих железо, кремнекислоту, гидроокиси алюминия и другие соединения, возможен ресинтез вторичных минералов (И. Н. Антипов-Каратаев, Е. И. Парфенова, Е. А. Ярилова). При ог-леении он наиболее проявляется в условиях ослабленного выноса продуктов глеевого процесса и почти не отмечается при хорошо выраженных нисходящих токах воды при кислой реакции почвенных растворов. Оно может происходить, по-видимому, в результате как ферментативной деятельности микроорганизмов, так и воздействия продуктов жизнедеятельности анаэробных микроорганизмов. К таким продуктам могут относиться газообразные соединения (Н2, Н25), низкомолекулярные органические кислоты и гуминовые кислоты.[ ...]
За счет выделения водорослями бактерицидных веществ происходит отмирание бактерий, и в частности патогенных кишечной группы. Поэтому в процессе доочистки сточных вод в биологических прудах имеет место не только удаление биогенных и органических веществ, но и бактериальных загрязнений. Как уже указывалось, для целей доочистки должны применяться строго аэробные биологические пруды. Обязательными условиями нормальной работы таких прудов является соблюдение оптимальных для водных организмов реакции среды (pH) и температуры, а также наличие растворенного кислорода не менее 1 мг/л. Важное значение имеет перемешивание воды, которое препятствует образованию анаэробных зон и способствует процессам стабилизации качества воды.[ ...]
О том же свидетельствует более длительное сохранение зеленой окраски у плодов, поскольку в условиях РГС подавляется распад хлорофилла. Снижение активности окислительных ферментов — полифенолоксидазы и аскор-бинатоксидазы, обусловленное уменьшением концентрации 02 в газовой среде, способствует лучшему сохранению Р- и С-витаминной активности плодов, препятствуя одновременно побурению последних. При концентрациях 02 и С02, рекомендуемых для вида и сорта плодов и овощей, в РГС наблюдается меньшее - накопление в их тканях ацетальдегида и спирта (продуктов анаэробного распада сахаров), что коррелирует с меньшим поражением плодов загаром. Существующая теория объясняет это явление так. В растительных тканях как при обычном содержании, так и при недостатке кислорода имеют место аэробный и анаэробный типы дыхания. В условиях, когда подавляется процесс аэробного дыхания (при понижении концентрации 02 в атмосфере), тормозится и дыхание анаэробное. Что же касается ацетальдегида, то его образование зависит и от реакции декарбоксилирования, а она, как уже отмечалось, в условиях РГС подавляется.[ ...]
Таким образом, исследования, проведенные на большинстве водохранилищ Волго-Камского каскада и ряде озер Латвии, показали, что наиболее общие, универсальные, интегрирующие деятельность всего бактериального сообщества показатели физиологической активности — интенсивность процессов темновой ассимиляции С02 и суммарная деструкция органического вещества — являются весьма информативными. Разнообразие характера грунтов обследованных водоемов позволило выявить влияние отдельных экологических факторов на интенсивность и направленность микробиологических процессов. Стало очевидным, что вблизи крупных городов существуют участки, где даже при достаточной аэрации грунтов в них превалируют анаэробные процессы, а общая активность бактериобентоса заметно снижаетея, несмотря на обеспеченность органическим веществом. Это — свидетельство постоянного антропогенного воздействия, и реакция микрофлоры на него является четким биологическим тестом. Удалось выявить, при каких условиях доминируют бактерии с гетеротрофным обменом и при каких — хемосинтезирующие, что может способствовать уточнению расчетов образования и деструкции органического вещества в водоемах.[ ...]
Значительно менее полно исследованы показатели углеводного обмена при адаптивной экзогенной гипоксии, т. е. при легком и умеренном дефиците кислорода в окружающей среде. Однако имеющиеся немногочисленные экспериментальные данные показывают, что и в этом случае происходят усиленное использование гликогена в мышцах, повышение содержания молочной кислоты и сахара в крови. Как и следовало ожидать, уровень насыщения воды кислородом, при котором отмечаются эти сдвиги, неодинаков для разных видов. Например, у миноги отмечена гипергликемия при снижении содержания кислорода всего лишь на 20% от исходного [379] и у 1 аЬео сарепвк концентрация сахара в крови оставалась постоянно низкой даже при 40% насыщения воды кислородом и только дальнейшее снижение насыщения приводило к быстрому увеличению уровня сахара в крови. Повышение сахара в крови и молочной кислоты отмечено при гипоксии у линя [238]. Аналогичная реакция на гипоксию отмечена и у канального сомика [275, 601]. В первой из этих работ при 50%-ном насыщении воды кислородом у рыб выявлено повышение содержания молочной кислоты, которое продолжалось и в первый час нормоксии, т. е. после возвращения рыб в нормальные кислородные условия. Восстановление биохимических показателей к норме происходило в течение 2—6 ч, а увеличение содержания лактата и дыхательного коэффициента с 0,8 до 2,0 свидетельствовало об увеличении анаэробного гликолиза.[ ...]
ru-ecology.info
Биохимия физической работы - Moveout
Прежде чем мы описать систему MOVEOUT, я хочу, чтобы вы вообще понимали какие процессы происходят в мышцах при работе. Я не буду вдаваться в мельчайшие подробности, дабы не травмировать вашу психику, поэтому расскажу о самом важном. Что же, возможно многие не поймут этот раздел, но советую его хорошо изучить, так как благодаря нему вы поймете как работают наши мышцы, а значит поймете как их правильно тренировать.
Итак, основное, что нужно для работы наших мышц – это молекулы АТФ с которой мышцы получают энергию. От расщепления АТФ образуется молекула АДФ + энергия. Вот только запасов АТФ хватает в наших мышцах всего на 2 секунды работы, а далее идет ресинтез АТФ из молекул АДФ. Собственно, от типов процессов ресинтеза АТФ и зависит работоспособность и функциональность.
Итак, выделяют такие процессы. Они обычно подключаются друг за другом
1. Анаэробный креатинфосфатный
Главным преимуществом креатинфосфатного пути образования АТФ являются
- малой время развертывания,
- высокая мощность.
Креатинфосфатный путь связан с веществом креатинфосфатом. Креатинфосфат состоит из вещества креатина. Креатинфосфат обладает большим запасом энергии и высоким сродством с АДФ. Поэтому он легко вступает во взаимодействие с молекулами АДФ, появляющимися в мышечных клетках при физической работе в результате реакции гидролиза АТФ. В ходе этой реакции остаток фосфорной кислоты с запасом энергии переносится с креатинфосфата на молекулу АДФ с образованием креатина и АТФ.
Креатинфосфат + АДФ → креатин + АТФ.
Эта реакция катализируется ферментом креатинкиназой. Данный путь ресинтеза АТФ иногда называют креатикиназным, иногда фосфатным или алактатным.
Креатинфосфат – вещество непрочное. Образование из него креатина происходит без участия ферментов. Не используемый организмом креатин, выводится из организма с мочой. Синтез креатинфосфата происходит во время отдыха из избытка АТФ. При мышечной работе умеренной мощности запасы креатинфосфата могут частично восстанавливаться. Запасы АТФ и креатинфосфата в мышцах называют также фосфагены.
Фосфатная система отличается очень быстрым ресинтезом АТФ из АДФ, однако она эффективна только в течение очень короткого времени. При максимальной нагрузке фосфатная система истощается в течение 10 с. Вначале в течение 2 с расходуется АТФ, а затем в течение 6-8 с - КФ.
Фосфатная система называется анаэробной, потому что в ресинтезе АТФ не участвует кислород, и алактатной, поскольку не образуется молочная кислота.
Эта реакция является главным источником энергии для упражнений максимальной мощности: бег на короткие дистанции, прыжки метания, подъем штанги. Эта реакция может неоднократно включаться во время выполнения физических упражнений, что делает возможным быстрое повышение мощности выполняемой работы.
2. Анаэробный гликолиз
По мере увеличения интенсивности нагрузки наступает период, когда мышечная работа уже не может поддерживаться за счет одной только анаэробной системы из-за нехватки кислорода. С этого момента в энергообеспечение физической работы вовлекается лактатный механизм ресинтеза АТФ, побочным продуктом которого является молочная кислота. При недостатке кислорода молочная кислота, образовавшаяся в первой фазе анаэробной реакции, не нейтрализуется полностью во второй фазе, в результате чего происходит ее накопление в работающих мышцах, что приводит к ацидозу, или закислению, мышц.
Гликолитический путь ресинтеза АТФ, так же как креатинфосфатный является анаэробным путем. Источником энергии, необходимой для ресинтеза АТФ в данном случае является мышечный гликоген. При анаэробном распаде гликогена от его молекулы под действием фермента фосфорилазы поочередно отщепляются концевые остатки глюкозы в форме глюкозо-1-фосфата. Далее молекулы глюезо-1-фосфата после ряда последовательных реакций превращаются в молочную кислоту. Этот процесс называется гликолиз. В результате гликолиза образуются промежуточные продукты, содержащие фосфатные группы, соединенные макроэргическими связями. Эта связь легко переносится на АДФ с образованием АТФ. В покое реакции гликолиза протекают медленно, но при мышечной работе его скорость может возрасти в 2000 раз, причем уже в предстартовом состоянии.
Время развертывания 20-30 секунд.
Время работы с максимальной мощностью – 2 -3 минуты.
Гликолитический способ образования АТФ имеет ряд преимуществ перед аэробным путем:
- он быстрее выходит на максимальную мощность,
- имеет более высокую величину максимальной мощности,
- не требует участия митохондрий и кислорода.
Однако у этого пути есть и свои недостатки:
- процесс малоэкономичен,
- накопление молочной кислоты в мышцах существенно нарушает их нормальное функционирование и способствует утомлению мышцы.
1. Аэробный путь ресинтеза
Аэробный путь ресинтеза АТФ иначе называется тканевым дыханием – это основной способ образования АТФ, протекающий в митохондриях мышечных клеток. В ходе тканевого дыхания от окисляемого вещества отнимаются два атома водорода и по дыхательной цепи передаются на молекулярный кислород, доставляемый в мышцы кровью, в результате чего возникает вода. За счет энергии, выделяющейся при образовании воды, происходит синтез молекул АТФ из АДФ и фосфорной кислоты. Обычно на каждую образовавшуюся молекулу воды приходится синтез трех молекул АТФ.
Кислородная, или аэробная, система является наиболее важной для спортсменов на выносливость, поскольку она может поддерживать физическую работу в течение длительного времени. Кислородная система обеспечивает организм, и в частности мышечную деятельность, энергией посредством химического взаимодействия пищевых веществ (главным образом, углеводов и жиров) с кислородом. Пищевые вещества поступают в организм с пищей и откладываются в его хранилищах для дальнейшего использования по необходимости. Углеводы (сахар и крахмалы) откладываются в печени и мышцах в виде гликогена. Запасы гликогена могут сильно варьироваться, но в большинстве случаев их хватает как минимум на 60-90 мин работы субмаксимальной интенсивности. В то же время запасы жиров в организме практически неисчерпаемы.
Углеводы являются более эффективным "топливом" по сравнению с жирами, так как при одинаковом потреблении энергии на их окисление требуется на 12% меньше кислорода. Поэтому в условиях нехватки кислорода при физических нагрузках энергообразование происходит в первую очередь за счет окисления углеводов.
Поскольку запасы углеводов ограничены, ограничена и возможность их использования в видах спорта на выносливость. После исчерпания запасов углеводов к энергообеспечению работы подключаются жиры, запасы которых позволяют выполнять очень длительную работу. Вклад жиров и углеводов в энергообеспечение нагрузки зависит от интенсивности упражнения и тренированности спортсмена. Чем выше интенсивность нагрузки, тем больше вклад углеводов в энергообразование. Но при одинаковой интенсивности аэробной нагрузки тренированный спортсмен будет использовать больше жиров и меньше углеводов по сравнению с неподготовленным человеком.
Таким образом, тренированный человек будет более экономично расходовать энергию, так как запасы углеводов в организме небезграничны.
Производительность кислородной системы зависит от количества кислорода, которое способен усвоить организм человека. Чем больше потребление кислорода во время выполнения длительной работы, тем выше аэробные способности. Под воздействием тренировок аэробные способности человека могут вырасти на 50%.
Время развертывания составляет 3 – 4 минуты, но у хорошо тренированных спортсменов может составлять 1 мин. Это связано с тем, что на доставку кислорода в митохондрии требуется перестройка практически всех систем организма.
Время работы с максимальной мощностью составляет десятки минут. Это дает возможность использовать данный путь при длительной работе мышц.
По сравнению с другими идущими в мышечных клетках процессами ресинтеза АТФ аэробный путь имеет ряд преимуществ:
- Экономичность: из одной молекулы гликогена образуется 39 молекул АТФ, при анаэробном гликолизе только 3 молекулы.
- Универсальность в качестве начальных субстратов здесь выступают разнообразные вещества: углеводы, жирные кислоты, кетоновые тела, аминокислоты.
- Очень большая продолжительность работы. В покое скорость аэробного ресинтеза АТФ может быть небольшой, но при физических нагрузках она может стать максимальной.
Однако есть и недостатки.
- Обязательное потребление кислорода, что ограничено скоростью доставки кислорода в мышцы и скоростью проникновения кислорода через мембрану митохондрий.
- Большое время развертывания.
- Небольшую по максимальной величине мощность.
Поэтому мышечная деятельность, свойственная большинству видов спорта, не может быть полностью получена этим путем ресинтеза АТФ.
Примечание. Эта глава написана на основе учебника "ОСНОВЫ БИОХИМИИ СПОРТА" Автор: Кучерявый Всеволод Владимирович.
moveout.in
Ресинтез АТФ — аэробный и анаэробный механизмы
АТФ в процессе сокращения поставляет необходимую энергию для образования актомиозинового комплекса, а в процессе расслабления мышцы — обеспечивает энергией активный транспорт ионов кальция в ретикулум. Для поддержания сократительной функции мышцы концентрация АТФ в ней должна находиться на постоянном уровне от 2 до 5 ммоль/кг.
Поэтому при мышечной деятельности аденозинтрифосфорная кислота должна восстанавливаться с той же скоростью, с какой расщепляется в процессе сокращения, что осуществляется отдельными биохимическими механизмами ее ресинтеза.
Энергетические источники ресинтеза АТФ в скелетных мышцах и других тканях — богатые энергией фосфатсодержащие вещества. Они присутствуют в тканях (креатинфосфат, аденозиндифосфат) или образуются в процессе катаболизма гликогена, жирных кислот и других энергетических субстратов. Кроме того, в результате аэробного окисления различных веществ возникают энергии протонного градиента на мембране митохондрий.
Ресинтез аденозинтрифосфата может осуществляться в реакциях без участия кислорода (анаэробные механизмы) или с его участием (аэробный механизм). В обычных условиях ресинтез АТФ в мышцах происходит преимущественно аэробным путем. При напряженной физической работе, когда доставка кислорода к мышцам затруднена, включаются и анаэробные механизмы ресинтеза АТФ. В скелетных мышцах человека выявлены три вида анаэробных и один путь аэробного восстановления аденозинтрифосфата.
К анаэробным механизмам относятся креатинфосфокиназный (фосфогенный или алактатный), гликолитический (лактатный) и миокиназный механизмы.
Аэробный механизм ресинтеза АТФ заключается в окислительном фосфорилировании, протекающем в митохондриях, количество которых в скелетных мышцах при аэробных тренировках существенно увеличивается. Энергетическими субстратами аэробного окисления служат: глюкоза, жирные кислоты, частично аминокислоты, а также промежуточные метаболиты гликолиза (молочная кислота) и окисления жирных кислот (кетоновые тела).
Каждый механизм имеет разные энергетические возможности, которые оцениваются по следующим критериям: максимальная мощность, скорость развертывания, метаболическая емкость и эффективность.
Максимальная мощность — это наибольшая скорость образования АТФ в данном метаболическом процессе. Она лимитирует предельную интенсивность работы, выполняемой за счет используемого механизма.
Скорость развертывания — время достижения максимальной мощности данного пути ресинтеза адено-зинтрифосфата от начала работы.
Метаболическая емкость — общее количество АТФ, которое может быть получено в используемом механизме ресинтеза АТФ за счет величины запасов энергетических субстратов. Емкость лимитирует объем выполняемой работы. Метаболическая эффективность — это та часть энергии, которая накапливается в макроэргических связях аденозинт-рифосфата. Она определяет экономичность выполняемой работы и оценивается общим значением коэффициента полезного действия, представляющего отношение всей полезно затраченной энергии к ее общему количеству, выделенному при текущем метаболическом процессе.
Общий коэффициент полезного действия при преобразовании энергии метаболических процессов в механическую работу зависит от двух показателей:
- эффективности фосфорилирования;
- эффективности хемомеханического сопряжения (эффективности преобразования АТФ в механическую работу).
Эффективность хемомеханического сопряжения в процессах аэробного и анаэробного метаболизма примерно одинакова и составляет 50%.
Эффективность фосфорилирования наивысшая в алактатном анаэробном процессе — около 80%, и наименьшая в анаэробном гликолизе — в среднем 44%. В аэробном же процессе она составляет примерно 60%.
Таким образом, анаэробные механизмы имеют большую максимальную мощность и эффективность образования АТФ, но короткое время удержания и небольшую емкость, из-за малых запасов энергетических субстратов. Например, максимальная мощность креатинфосфокиназной реакции развивается уже на 0,5—0,7 с интенсивной работы и поддерживается 10-15 с у нетренированных людей идо 25-30 су высокотренированных спортсменов и составляет 3,8 кДж/кг в минуту.
Гликолитический механизм ресинтеза АТФ отличается невысокой эффективностью. Большая часть энергии остается в молекулах образующейся молочной кислоты. Концентрация последней находится в прямой зависимости от мощности и продолжительности работы, и может быть выделена только путем аэробного окисления.
Гликолиз — это основной путь энергообразования в упражнениях субмаксимальной мощности, предельная продолжительность которых составляет от 30 с до 2,5 мин (бег на средние дистанции, плавание на 100 и 200 м и др.).
Гликолитический механизм энергообразования служит биохимической основой специальной скоростной выносливости организма.
Миокиназная реакция происходит в мышцах при значительном увеличении концентрации АДФ в саркоплазме. Такая ситуация возникает при выраженном мышечном утомлении, когда другие пути ресинтеза уже не возможны.
Таким образом, анаэробные механизмы являются основными в энергообеспечении кратковременных упражнений высокой интенсивности.
При адаптации к интенсивным нагрузкам повышается активность ферментов анаэробных механизмов и запасов энергетических механизмов: содержание креатинфосфата в скелетных мышцах может увеличиваться в 1,5—2 раза, а содержание гликогена — почти в 3 раза.
www.fizkulturaisport.ru








